elocation-id: e4239
In recent years, the macroalga Sargassum spp. has become an economic and environmental problem in several coastal regions. Nonetheless, due to the wide variety of bioactive compounds it possesses, it has been proposed as a sustainable alternative in agriculture, as it can improve seed germination, nutrient absorption, and photosynthesis, and mitigate biotic and abiotic stress. The objective of this study was to evaluate the effect of priming tomato seeds with aqueous extracts of Sargassum spp. on germination, vigor, biomass, photosynthetic pigments and some indicators of the antioxidant system of seedlings. The concentrations of the extracts were: 0.5, 1.5, 2.5 and 3.5%, and a control with distilled water. The results indicate that the extracts improved germination, vigor, phenolic compounds, flavonoids, antioxidant capacity and photosynthetic pigments. There were no significant improvements in biomass. These results suggest that applying Sargassum spp. extracts may be an ecological alternative to improve germination parameters and stimulate the synthesis of secondary metabolites in tomato seedlings.
Solanum lycopersicum L., biostimulants, seed priming
Modern agriculture faces the challenge of increasing crop productivity and quality under criteria of sustainability and food security (FAO, 2025). The growing demand for food drives strategies that optimize agricultural yield while reducing environmental impact. In this context, natural biostimulants stand out for their capacity to improve production and quality, decreasing the use of agrochemicals and mitigating biotic and abiotic stress (Durán-Hernández et al., 2022). Among them, seaweed derivatives, such as Sargassum spp., are promising due to their wealth of bioactive compounds. These macroalgae contain polysaccharides, phenolic compounds, carotenoids, phytohormones and amino acids, which improve germination, growth and quality, as well as confer tolerance to salt, water, thermal and pathogen stress (Senthilkumar et al., 2024).
Although its massive proliferation generates environmental and economic problems in the Mexican Caribbean, its high content of secondary metabolites makes it a valuable source of agricultural biostimulants (Adderley et al., 2023). Previous studies show its potential to improve the growth and nutraceutical quality of different crops (Rivera-Solís et al., 2021).
For its part, the tomato (Solanum lycopersicum L.) is one of the most cultivated and consumed vegetables worldwide, appreciated for its nutritional and antioxidant value (Collins et al., 2022). To ensure the productivity of this crop, high-quality seeds are required to ensure uniform germination, a key stage for crop yield (Ruiz-Ramirez et al., 2021; Reed et al., 2022). Germination, the transition from dormancy to active growth, can be optimized by pre-germination treatments such as priming, which improves the germination rate, vigor and stress tolerance (Wang and Shi, 2024; Abinandan et al., 2025). Given the biostimulant potential of Sargassum spp., this study evaluated the effect of Sargassum spp. extracts on germination, photosynthetic pigments and antioxidant activity in tomato seedlings.
Brown algae Sargassum spp., previously dehydrated and pulverized, provided by the Antonio Narro Agrarian Autonomous University (UAAAN), was used. The liquid extract was prepared according to the methodology of Sariñana-Aldaco et al. (2021), where its biochemical characterization is detailed. A mother solution (100%) was prepared in a 1:20 ratio (m:v, algae:distilled water), boiled for 1 h, pressed and filtered. From this concentrated solution, the required dilutions (0.5, 1.5, 2.5 and 3.5%) were obtained with bidistilled water.
Saladette tomato seeds of the ‘Río Grande’ variety were used, disinfected with 75% ethanol for 5 min and washed twice with distilled water. The seeds were soaked for 24 h in the concentrations of the Sargassum spp. extract (0.5, 1.5, 2.5 and 3.5%), including a control with distilled water. Subsequently, they were placed on #1 filter paper inside sterile Petri dishes (90 mm), with 10 seeds per dish and 20 dishes per treatment. The filter paper was moistened with sterile distilled water. The experimental design was completely randomized with five treatments and 20 repetitions. The Petri dishes were incubated in a germination chamber (Achieva, Lab-Tech Inc, Model No. A-3920), under a 12-hour day/night cycle, at 25 ±2 °C and 60% relative humidity (Li et al., 2019). After 14 days, the germination percentage, fresh biomass and vigor percentage were evaluated.
Germination percentage (G) and vigor (V) were calculated using the following formulas:
Where: n= number of germinated seeds on day 14; N= total number of seeds.
Where: Pn= normal seedlings; N= total number of seeds.
Chlorophyll content was determined according to the method of Lichtenthaler and Wellburn (1983), using 95% ethanol as a solvent. Absorbance readings were performed at 665, 649, and 470 nm in a UV-visible spectrophotometer (Jenway 7305).
Chlorophyll concentrations were calculated using the following equations:
A quantity of 2 g of fresh sample was homogenized with 10 ml of 80% ethanol, by constant orbital agitation (70 rpm) for 24 h at 70 rpm and 5 °C. Subsequently, the extracts were centrifuged at 3 000 rpm for 5 min, and the supernatant was used for subsequent analyses.
The content of phenolic compounds was determined using an adaptation of the Folin-Ciocalteu method (Singleton et al., 1999). Fifty µl of ethanolic extract was mixed with 3 ml of distilled water and 250 µl of Folin-Ciocalteu reagent (1 N). After 3 minutes of rest, 750 µl of Na2CO3 and 950 µl of distilled water were added, incubating for 2 h. Absorbance was measured at 765 nm in a UV-visible spectrophotometer (Jenway 7305), expressing the results as mg of gallic acid equivalents (GA) per 100 g of fresh weight (FW).
Total flavonoid content was quantified using the colorimetric method described by Buendía-García et al. (2021). 250 µl of extract were mixed with 1.25 ml of distilled water and 75 µl of NaNO2 (5%), resting for 5 min. Then, 150 µl of AlCl3, 500 µl of NaOH (1 M), and 275 µl of distilled water were added. Absorbance was measured at 510 nm, and the results were expressed in mg of quercetin equivalents (QE) per 100 g FW.
Antioxidant capacity was evaluated using the DPPH+ method (Brand-Williams et al., 1995), using a solution of DPPH+ in ethanol (0.025 mg ml-1). A quantity of 50 µl was mixed with 1 950 µl of DPPH+, incubated for 30 min, and absorbance was measured at 517 nm. The results were expressed in milliequivalents (Meq) of Trolox per 100 g of FW.
Catalase activity (CAT, EC 1.11.1.6) was determined according to the method of Aebi (1974). Readings were performed at room temperature in a UV-visible spectrophotometer (Jenway 7305), monitoring the decrease in absorbance at 240 nm due to H2O2 decomposition. Enzymatic activity was calculated using the molar extinction coefficient (ε240 = 43.6 M cm-1) and protein concentration, determined by the method of Bradford (1976). Results were expressed as U mg-1 protein, defining one unit (U) as the amount of enzyme that causes a 0.001 absorbance change per minute under the assay conditions.
Data normality was verified using the Kolmogorov-Smirnov test. Data expressed as a percentage were transformed using arcsine or square root functions, depending on their distribution. The analysis of variance and the comparison of means by Tukey’s test (p ≤ 0.05) were performed with the Statistical Analysis System (SAS) statistical package, version 9.3.
Germination and vigor are critical stages in plant development, determining crop establishment and yield (Papoui et al., 2025). In this study, treatment with the liquid extract of Sargassum spp. significantly improved the germination and vigor of tomato seeds (Figure 1). The 3.5% concentration increased germination by 19%, while 1.5% increased vigor by 13% compared to the control.
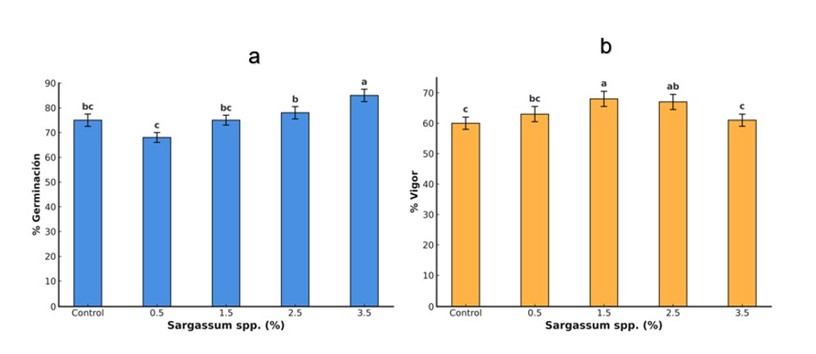
Similar results have been reported in tomatoes with brown algae extracts (Hernández-Herrera et al., 2023) and in Abelmoschus esculentus with Ecklonia maxima extracts (Makhaye et al., 2021). The positive effect is attributed to the presence of phytohormones, polysaccharides and antioxidant compounds that regulate key processes during germination (Cossa et al., 2023; Margal et al., 2023). Brown algae, which contain auxins, gibberellins and cytokinins that facilitate the breaking of dormancy by activating hydrolytic enzymes, such as amylase, which converts starch into simple sugars for embryonic development (Sariñana-Aldaco et al., 2022). In addition, these phytohormones promote cell division and elongation (Zluhan-Martínez et al., 2021). Nevertheless, its effectiveness depends on multiple factors, such as the species of algae, type of extraction, environmental conditions, concentration and the receiving plant species (Martínez-González et al., 2022). High concentrations can cause adverse effects due to the accumulation of compounds that alter the hormonal balance or induce physiological stress (Castro et al., 2022).
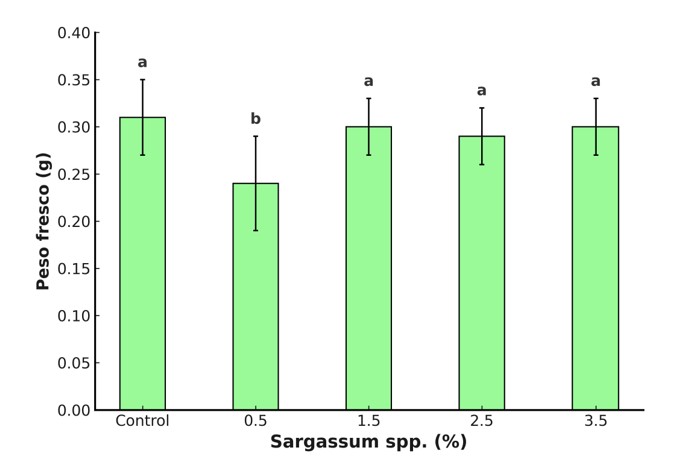
Priming with liquid extract of Sargassum spp. did not cause significant effects on biomass (Figure 2). These results agree with previous studies that report no differences between different concentrations used. For example, Morales-Meléndez et al. (2023) observed no significant increases in biomass production of tomato plants treated with seaweed extracts, and Rivera-Solís et al. (2021) found no differences compared to the control. This suggests that the chemical composition of the extracts and their concentration are key factors in inducing a positive physiological response or preventing inhibitory effects.
Total chlorophyll and carotenoids are key indicators of the photosynthetic capacity and physiological state of plants (Sherin et al., 2022). In this study, both pigments were significantly affected by Sargassum spp. extracts (Table 1). The 3.5% concentration reduced total chlorophyll by 14.85% compared to the control. Similar results have been reported with seaweed extracts (Salazar-Salazar et al., 2022; Mohammed et al., 2023).
| Sargassum spp. extract (%) | Total chlorophyll (mg g-1 DW) | Carotenoids (mg g-1 DW) |
|---|---|---|
| Control | 1.28 b | 102.68 b |
| 0.5 | 1.48 ab | 118.27 ab |
| 1.5 | 1.91 a | 137.8 a |
| 2.5 | 1.61 a | 128.44 ab |
| 3.5 | 1.09 b | 82.91 b |
This effect has been attributed to the action of betaines present in the extracts, which act as molecular chaperones stabilizing sensitive biomolecules, such as pigments, and preventing their degradation (Sariñana-Aldaco et al., 2025). Such compounds have been shown to prevent the loss of photosynthetic activity by inhibiting chlorophyll degradation (Genard et al., 1991), especially at intermediate doses.
The plant antioxidant system comprises two components: enzymatic, considered the first line of defense, integrated by enzymes such as catalase (CAT), ascorbate peroxidase (APX) and glutathione peroxidase (GPX), which degrade reactive oxygen species (ROS) such as H2O2 (Dumanović et al., 2021), and non-enzymatic, made up of antioxidant metabolites such as ascorbic acid, phenolic compounds, glutathione and carotenoids (Hashim et al., 2020). In this study, the Sargassum spp. extract (1.5%) significantly increased total phenols (141%), flavonoids (125%), and antioxidant capacity (11%) compared to the control (Figure 3). However, CAT activity decreased in treatments with 1.5% and 3.5% (Figure 4). These results agree with reports on the positive effect of seaweed extracts on the accumulation of bioactive compounds (Sariñana-Aldaco et al., 2021); nevertheless, they differ from studies that report increases in antioxidant enzymes in soybeans treated with Ascophyllum nodosum extracts (Repke et al., 2022).
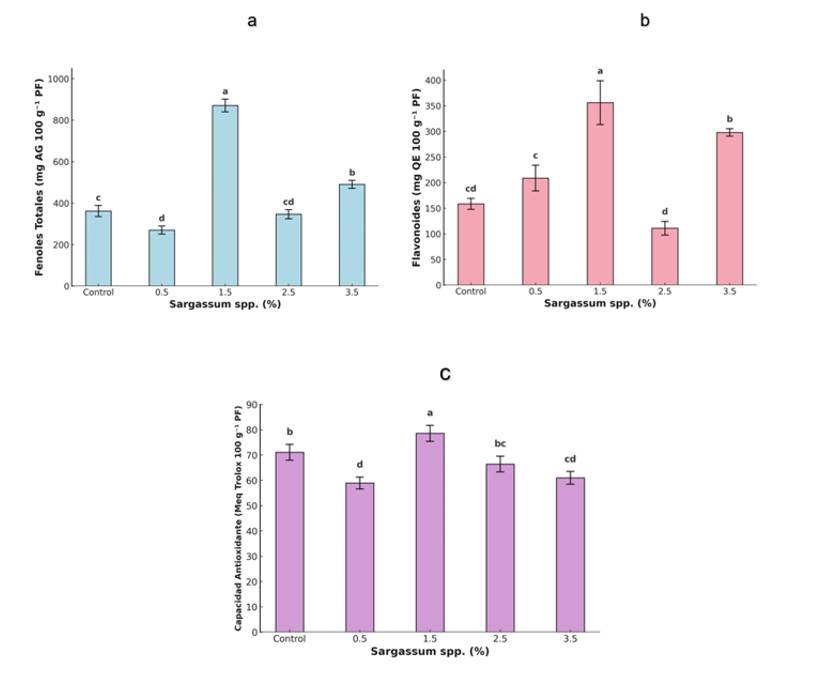
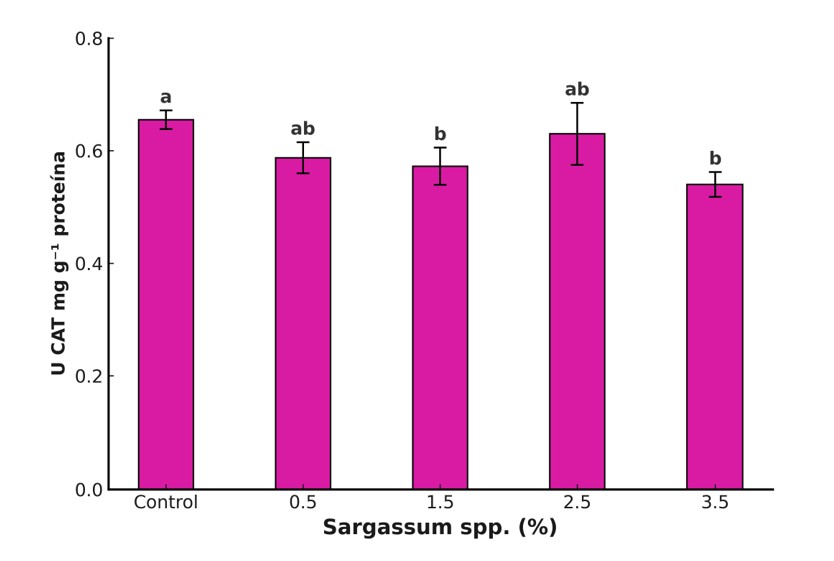
The observed response can be attributed to metabolites of the extract that activate signaling and gene expression cascades for antioxidant enzymes and synthesis of phenolic compounds (González-Morales et al., 2021). In addition, some metabolites exert direct action on the cytoplasm (Lau et al., 2025). The reduction in CAT suggests a rebalancing of the antioxidant system, favoring non-enzymatic defense through the biosynthesis of phenols and flavonoids, which decreases the need for enzymatic pathways to neutralize ROS (Kumar et al., 2024).
Sargassum spp. extract, as a natural biostimulant, improved tomato seed germination, seedling vigor and the biosynthesis of bioactive compounds, thereby increasing their antioxidant capacity. Sargassum spp. extract is a sustainable alternative to promote germination, seedling development, and increased production of secondary metabolites.
Adderley, A.; Wallace, S.; Stubbs, D.; Bowen-O’Connor, C.; Ferguson, J.; Watson, C. and Gustave, W. 2023. Sargassum sp. as a biofertilizer: is it really a key towards sustainable agriculture for The Bahamas? Bulletin of the National Research Centre. 47(1):1-11. https://doi.org/10.1186/s42269-023-01087-w.
Buendía-García, A.; Lozano-Cavazos, C. J.; Rodríguez-Ortiz, J. C.; Carballo-Méndez, F. D. J.; Moreno-Reséndez, A.; Sariñana-Navarrete, M. D. A. y Preciado-Rangel, P. 2021. La biofortificación con selenio mejora la calidad nutracéutica y la capacidad antioxidante de la lechuga (Lactuca sativa L.). Ecosistemas y Recursos Agropecuarios. 8(3):1-8. https://doi.org/10.19136/era.a8n3.3104.
Hernández-Herrera, R. M.; González-González, M. F.; Velasco-Ramírez, A. P.; Velasco-Ramírez, S. F.; Santacruz-Ruvalcaba, F. and Zamora-Natera, J. F. 2023. Seaweed extract components are correlated with the seeds germination and growth of tomato seedlings. Seeds. 2(4):436-448. https://doi.org/10.3390/seeds2040033.
Mohammed, S.; El-Sheekh, M. M.; Aly, S. H.; Al-Harbi, M.; Elkelish, A. and Nagah, A. 2023. Inductive role of the brown alga Sargassum polycystum on growth and biosynthesis of imperative metabolites and antioxidants of two crop plants. Frontiers in Plant Science. 14(11):1-12. https://doi.org/10.3389/fpls.2023.1136325.
Morales-Meléndez, R.; Betancourt-Galindo, R.; Juárez-Maldonado, A.; Hernández-Pérez, A.; González-Fuentes, J. A.; Puente-Urbina, B. y Méndez-López, A. 2023. Aplicación de extractos de algas, NP’SZnO y microorganismos sobre la biomasa vegetal en tomate. Ecosistemas y Recursos Agropecuarios. 10(2):1-11. https://doi.org/10.19136/era.a10n2.3206.
Rivera-Solís, L. L.; Rodríguez-Jasso, R. M.; Flores-López, M. L.; Robledo-Olivo, A.; Sandoval-Rangel, A.; Sariñana-Aldaco, O. y González-Morales, S. 2021. Extractos de Sargassum spp. como inductores de tolerancia a Fusarium oxysporum en plántulas de tomate. Ecosistemas y Recursos Agropecuarios. 8(1):1-8. https://doi.org/10.19136/era.a8n1.2826.
Sariñana-Aldaco, O.; Benavides-Mendoza, A.; Juarez-Maldonado, A.; Robledo-Olivo, A.; Rodríguez-Jasso, R. M.; Preciado-Rangel, P. y Gonzalez-Morales, S. 2021. Efecto de extractos de Sargassum spp. en el crecimiento y antioxidantes de plántulas de tomate. Ecosistemas y Recursos Agropecuarios. 8(2):1-15. https://doi.org/10.19136/era.a8n2.2814.
Sariñana-Aldaco, O.; Rivera-Solís, L. L.; Benavides-Mendoza, A.; Robledo-Olivo, A., Rodríguez-Jasso, R. M. and González-Morales, S. 2025. Using brown algae in the plant-soil system: a sustainable approach to improving the yield and quality of agricultural crops. Horticulturae. 11(1):1-29. https://doi.org/10.3390/horticulturae11010094.
Senthilkumar, S.; Kuppusamy, S.; Palai, S.; Rajanbabu, V. and Vennila, A. 2024. Exploring Sargassum extract as a bio stimulant: advancements and benefits in soil plant systems a review. Communications in Soil Science and Plant Analysis. 55(22):3638-3648. https://doi.org/10.1080/00103624.2024.2397013.
Zluhan-Martínez, E.; López-Ruíz, B. A.; García-Gómez, M. L.; García-Ponce, B.; De la Paz Sánchez, M.; Álvarez-Buylla, E. R. and Garay-Arroyo, A. 2021. Integrative roles of phytohormones on cell proliferation, elongation and differentiation in the Arabidopsis thaliana primary root. Frontiers in Plant Science. 12(6):1-20. https://doi.org/10.3389/fpls.2021.659155.