elocation-id: elocation-id: e4327
The rehabilitation of degraded soils in semi-arid areas requires woody species with high water-stress tolerance and the potential to provide ecosystem services. Prunus serotina has been characterized as tolerant to water stress and having low soil requirements, in addition to presenting rapid growth; however, there is little recent evidence on its initial performance in semi-arid conditions in Mexico. The objective was to evaluate the early adaptation of P. serotina in the field and to estimate its regional environmental suitability. The study was conducted from 2021 to 2022 in Nopaltepec (Tula River Basin, State of Mexico, Mexico). A plantation trial was established with survival and growth monitoring; survival was analyzed using the Kaplan-Meier estimator and factors associated with mortality risk were analyzed using a Cox proportional hazards model. Environmental suitability was modeled with MaxEnt using bioclimatic and topographic variables; performance was verified using AUC and TSS and a suitability map was generated. Survival at 12 months was ≈68.3% and final diameter was associated with a lower risk of mortality (HR< 1), suggesting that a greater allocation to structural reserves improves stress tolerance. In the SDMs, annual precipitation, thermal seasonality and altitude contributed significantly, delimiting areas with high suitability in and around the basin. It was concluded that P. serotina is a viable option for restoration initiatives in semi-arid environments, recommending selection based on initial size, protection against mechanical damage and drought, and establishment in favorable climatic windows.
Prunus serotina, habitat suitability, mortality risk, survival.
The global environmental crisis, intensified by urbanization, agricultural expansion, land-use changes and the introduction of invasive exotic species, has accelerated natural degradation processes and compromised the resilience of ecosystems (Davies et al., 2012). This deterioration is manifested in a socioecological crisis with local and global effects, reflected in the loss of biodiversity, the reduction of ecosystem services and the decrease in the productivity of natural systems (Challenger and Dirzo, 2009).
In Mexico, the loss of natural cover has been particularly significant. In 1976, about 80% of the territory was covered by natural vegetation, while by 2011, this area had decreased to 71.7% (140 million hectares); by contrast, the areas devoted to agriculture, urbanization, and other anthropogenic uses increased to 28% (55 million hectares) (SEMARNAT, 2015). This trend is related to inadequate land use policies and poor territorial planning, which have accelerated degradation processes with potentially irreversible effects (Gardi et al., 2014).
Soil degradation is one of the most severe environmental problems. Factors such as overgrazing, population pressure, poverty and inadequate land management accelerate the loss of its biological productivity (Gardi et al., 2014). In 2002, approximately 44.9% of Mexican territory already showed signs of degradation (SEMARNAT and CP, 2003). Faced with this situation, there has been a promotion of production alternatives aimed at the rehabilitation of soils and ecosystems, among which agroforestry stands out, which integrates perennial woody species with crops or livestock, favoring ecological recovery and sustainable production (FAO, 2017).
The selection of species suitable for rehabilitation is crucial, as they must tolerate local conditions and provide ecological and productive benefits. Their incorporation can be assessed using species distribution models (SDMs), such as MaxEnt, which predict potential distribution based on presence data and environmental variables (Phillips et al., 2006). These tools, combined with survival analyses and Cox’s proportional hazard models, enable us to estimate the probability of persistence and the risk of mortality of species under specific conditions (Cox, 1972; Sigala et al., 2015).
In this context, Prunus serotina (P. serotina), commonly known as capulín, is emerging as a species with high potential for soil rehabilitation in semi-arid temperate regions of Mexico, thanks to its nutritional and cultural value, its tolerance to droughts and low temperatures, and its ability to grow in poor soils (Raya-Pérez et al., 2012). This study aimed to evaluate the adaptability of P. serotina in a degraded area of semi-arid temperate climate in the Tula micro-basin, Mexico, through the integration of species distribution models and field survival and growth analyses, in order to propose its incorporation into agroforestry arrangements aimed at the rehabilitation of degraded soils in semi-arid temperate regions of Mexico.
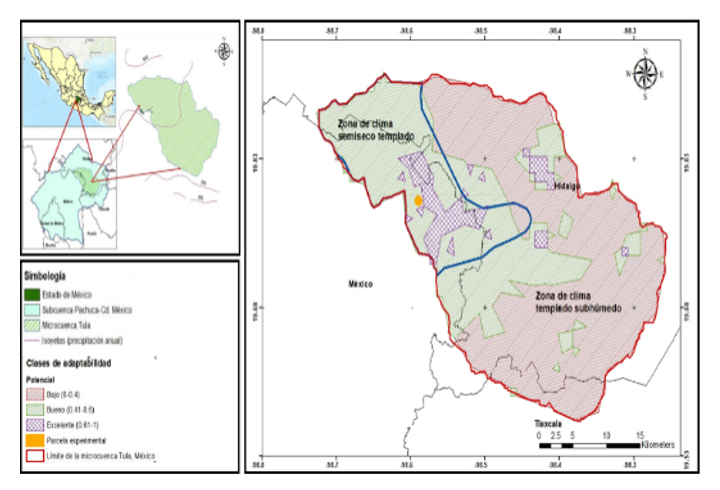
The study was conducted in the Tula micro-basin, located between the states of Mexico, Hidalgo and Tlaxcala, with an approximate area of 1 037.66 km² and altitudes ranging between 2 333 and 3 223 m. The average annual rainfall ranges from 500 to 700 mm, and the temperature ranges from 13 to 15 °C (INEGI, 2008). The predominant vegetation corresponds to alligator juniper, pine and oak forests, as well as succulent-stem xerophytic scrubland (INEGI, 2017). Two types of climate are distinguished: semi-dry temperate with rainfall in summer (28% of the surface) and Phaeozems haplic and Cambisols eutrophic; and subhumid temperate (72%) with Phaeozems haplic and Vertisol pelic (INEGI, 2017).
The experimental site was established in an area of 1 559 m², located in the ejido of the community of San Felipe Teotitlán, municipality of Nopaltepec, within the semi-dry zone of the micro-basin. The climate has an average annual temperature between 15 and 16.9 °C. The total annual rainfall ranges from 558.6 to 600 mm; the area depends on rainfed agriculture and shows soil degradation processes associated with overgrazing, logging, and land-use change (PMDU, 2019).
The field experiment was implemented under an agroforestry arrangement of boundary trees, with a distance of 5 m between individuals (Figure 1). The orientation of the rows was determined following the direction of the slope in order to reduce surface runoff and sediment loss and to favor the creation of local microclimates (CONABIO, 2011). Planting holes measuring 30 × 30 × 40 cm were excavated, where 41 seedlings were transplanted in July 2021.
Monitoring was carried out for 12 months (July 2021-June 2022). The following variables were recorded at each monthly evaluation: survival (1= live, 0= dead), total height (cm) with a tape measure, and stem diameter (cm) with a digital vernier caliper. Each individual (n= 41) was considered a unit of analysis.
The distribution model of P. serotina was constructed based on presence records obtained from GBIF (2022) and a set of 19 bioclimatic variables extracted from the WorldClim v.2 platform (Fick and Hijmans, 2017) in raster format with a spatial resolution of 30 arcseconds (~1 km²). Variables included: mean annual temperature (BIO1), mean diurnal range (BIO2), isothermality (BIO3), temperature seasonality (BIO4), maximum temperature of the hottest month (BIO5), minimum temperature of the coldest month (BIO6), annual temperature range (BIO7), mean temperature of the wettest quarter (BIO8), mean temperature of the driest quarter (BIO9), mean temperature of the warmest quarter (BIO10), mean temperature of the coldest quarter (BIO11), annual precipitation (BIO12), precipitation of the wettest month (BIO13), precipitation of the driest month (BIO14), seasonality of precipitation (BIO15), precipitation of the wettest quarter (BIO16), precipitation of the driest quarter (BIO17), precipitation of the warmest quarter (BIO18), and precipitation of the coldest quarter (BIO19). In addition, the elevation variable, obtained from the same platform with the same spatial resolution, was used. All variables were standardized and processed in raster format.
Multicollinearity was controlled by linear correlation (r≥ 0.8) in R (R Core Team, 2018). Modeling was performed with MaxEnt v.3.4.1 (Phillips et al., 2017). The algorithm configuration was: use of the random seed option so that a different set of test points were included in each repetition, 70% of the data was used for training and 30% for bootstrap resampling tests, 100 repetitions with logistic format and a maximum number of interactions equal to 1 000 were used, following methodological recommendations from Phillips et al. (2006) and Plasencia-Vázquez et al. (2014).
The predictive capacity of the models was evaluated using the area under the curve (AUC) of the ROC curve (Phillips and Dudík, 2008); employing 100 replicates, the median of AUC values was used to reduce the influence of extreme values (Plasencia-Vázquez et al., 2014). According to the classification proposed by Peterson et al. (2011), models with AUC> 0.9 have high predictive capacity, between 0.6 and 0.9 good capacity and <0.5 predictive capacity lower than expected by chance. The contribution of the variables to the model was determined using the Jackknife test (Phillips et al., 2006). The results of the MaxEnt model were exported to ArcGIS, where two mapping products were generated: i) a continuous habitat suitability map; and ii) a binary presence-absence map. Suitability thresholds were established following Qin et al. (2017), where values above 0.6 indicate high planting potential, values between 0.4 and 0.6 indicate good planting potential and values below 0.4 indicate low planting potential.
The survival, growth, and mortality risks of P. serotina were evaluated based on morphological variables measured in the field.
Estimation of survival. The survival rate was estimated using the Kaplan-Meier estimator, considering complete and censored observations (Kaplan and Meier, 1958). Survival was classified into four ranges: excellent (≥90%), acceptable (70-89%), marginal (50-69%), and not acceptable (<49%) (Rodríguez-Echeverry and Leiton, 2020).
Estimation of the risk of death. The risk of death was assessed using Cox’s proportional hazards regression (Cox, 1972); it was used to estimate the effect of the planting site and the production system on the morphological variables of the plant as covariates. The analysis was performed using SPSS v25 software.
Estimation of vegetative growth. The height growth rate (HGR, cm month-1) and the diameter growth rate (DGR, mm month-1) were calculated using equations (Griscom et al., 2005), organized in quarterly intervals corresponding to the seasons of the year.
In the Tula micro-basin, eight records of the presence of P. serotina were identified; nevertheless, in the surrounding areas, an additional 156 records were detected. Since the micro-basin is part of the Pachuca-Mexico City basin, the modeling was carried out for the entire area (164 records in total), and later the results were limited to the Tula micro-basin using ArcGIS.
The model presented high predictive capacity, with AUC values of 0.905 (training) and 0.916 (testing). After correlation analysis and variance inflation factor (VIF) calculations, six non-collinear variables were selected for modeling in MaxEnt: BIO1, BIO2, BIO6, BIO7, BIO12, and BIO15. The Jackknife test showed that three variables explained 74.9% of the total contribution of the model: BIO1 (42.3%), BIO12 (16.9%) and BIO7 (15.7%). The remaining ones contributed 25.1%: BIO15 (9.3%), BIO6 (8.1%) and BIO2 (7.6%).
Figure 2 shows the potential distribution of P. serotina in the micro-basin. The results indicate clear differences between climatic zones: a) semi-dry temperate area: the good category (0.41-0.6) predominated with 78.9% of the area, followed by the high category (0.61-1) with 17.1% and the low category (0-0.4) with 4%; and b) subhumid temperate area: the low category (0-0.4) predominated with 73.5%, followed by the good category (0.41-0.6) with 25.2% and the high category (0.61-1) with 1.3%.
Of the 41 individuals of P. serotina established in the experimental plot, a survival rate of 68.3% was obtained at the end of the 12-month evaluation period (July 2021-June 2022). During the first four months of evaluation, 100% survival was maintained, then it began to decrease until reaching the aforementioned percentage. For individuals who did not die (censored), the cumulative risk of survival increased from month 4 (October) to month 7 (January) by 15% and concluded at 22% at month 12 (June).
Cox’s model was significant (Chi²= 19.626, p = 0.003), confirming covariates influencing survival. The final diameter showed a positive effect: each 1 cm increase in diameter totally reduces the risk of death in P. serotina, controlling for other variables (Table 1).
Regarding the mean growth rates of plant height and stem diameter for the species during the summer cycle, P. serotina showed positive growth rates in height and diameter, with 1.033 cm month-1 and 0.201 cm month-1, respectively. During the second period, which corresponds to the autumn cycle, the species exhibited positive growth rates in height and stem diameter, with 0.134 cm month-1 and 0.176 cm month-1, respectively. In the winter cycle, which corresponds to the third period, the species showed positive growth rates for height and stem diameter, with 0.201 cm month-1 and 0.135 cm month-1, respectively. During the fourth period, which corresponds to the spring cycle, the species showed no changes in growth rates in both height (0 cm month-1) and stem diameter (-0.007 cm month-1). The species P. serotina showed the greatest reduction in height and diameter in this period compared to the previous three seasonal cycles.
The distribution model indicated that the semi-dry areas of the basin had the highest planting potential, which is consistent with the survival rate obtained in the experimental site (68.3%), considered marginally acceptable. These sites are characterized by average temperatures of 14 °C, annual rainfall of 500-600 mm, and Phaeozems haplic and Cambisols eutrophic (INEGI, 2008), conditions that correlate with the species’ natural habitats. Planting potential was strongly associated with climatic variables, such as BIO1 and BIO12, and seasonality between them, confirming that precipitation regimes significantly influence survival (Hernández et al club., 2010).
The observed survival rate coincides with that indicated by Navarro et al. (2006), who highlight that the establishment stage is critical in reforestation, and with that reported by Vallejo et al. (2012) and Ortega et al. (2006), who attribute survival to abiotic, biotic, and anthropic factors. During the first three months, P. serotina showed positive growth in height and diameter, associated with the rainy season, in agreement with the results of Pimienta-Barrios et al. (2002) and Pineda-Herrera et al. (2015), who also recorded greater growth under wet conditions.
In autumn and winter, growth decreased due to low temperatures and reduced humidity, which caused physiological dormancy and accumulation of reserves in stems and roots, as described by Díaz-Montenegro (2002) and Viveros and Vargas (2007). At the end of winter, growth stopped; however, with the arrival of spring and the increase in temperature and precipitation, budding was reactivated, a pattern similar to that reported by Pineda-Herrera et al. (2015) in forest species of Oaxaca.
The decrease in growth rate was more evident in height than in diameter because, in autumn and winter, low temperatures (12-13 °C, minimum 5-6 °C) and lower environmental humidity favored leaf senescence and physiological dormancy, associated with the reduction of auxins, gibberellins and cytokinins, and the increase in abscisic acid (Díaz-Montenegro, 2002). As an adaptive response to water stress and nutrient scarcity, accumulation of reserves in stems and roots was observed due to nutrient translocation (Díaz-Montenegro, 2002). At the end of winter, no growth was recorded, which is explained by the fact that deciduous species concentrate energy on breaking dormancy and initiating bud break (Viveros and Vargas, 2007).
With the arrival of spring, the increase in temperature (17 °C) and precipitation (>50 mm) favored the reactivation of growth hormones and the transport of photosynthates, thereby generating conditions opposite to those of winter conditions (Díaz-Montenegro, 2002; Villar et al., 2008). Similar results were reported by Pineda-Herrera et al. (2015) for forest species of Oaxaca, where they identified a minimum phase of growth between October and May, followed by an increase with the beginning of the rainy season. Individuals with anthropic damage showed reductions in growth, less marked in diameter, which coincides with Villar et al. (2008), who point out that stems and roots store carbohydrates that sustain development under stress. Finally, Cox’s risk analysis confirmed that the final diameter significantly reduces the risk of mortality, which supports the hypothesis of Fontes (1999), who points out that some species allocate more resources to diameter increase in early stages as an adaptive strategy to site conditions.
The study demonstrated that integrating species distribution models with analyses of survival, vegetative growth and mortality risk is an effective tool for assessing the adaptability of multipurpose species in degraded soil rehabilitation contexts. The species P. serotina exhibited an adequate planting potential under the soil and climatic conditions of the Tula micro-basin, as shown by the distribution model. The survival rate in the field was marginally acceptable, which highlights the need to consider seasonal factors in its management.
Vegetative growth was closely linked to climatic seasonality: during the rainy season, there was a positive growth in height and diameter, whereas in autumn and winter, low temperatures and decreased humidity promoted leaf senescence and physiological dormancy, with the accumulation of reserves in stems and roots. In spring, the absence of observable growth was associated with the use of these reserves to break dormancy and initiate regrowth. Regarding the risk of mortality, morphological variables were decisive. It is worth highlighting that the final stem diameter reduced the risk of death by 100% for each centimeter of increase, assuming the other variables remained constant, which confirms its relevance as an indicator of early adaptation to the site.
Davies, J.; Poulsen, L.; Schulte-Herbrüggen, B.; Mackinnon, K.; Crawhall, N.; Henwood, W. D.; Dudley, N.; Smith, J. and Gudka, M. 2012. Conserving dryland biodiversity. International union for conservation of nature, united nations environment programme-world conservation monitoring programme y united nations convention to combat desertification. Gland, Suiza. 84 p. https://portals.iucn.org/library/sites/library/files/documents/2012-050.pdf.
Gardi, C.; Angelini, M. E.; Barceló, S.; Comerma, J. A.; Cruz-Gaistardo, C.; Encina-Rojas, A.; Jones, A.; Krasilnikov, P.; Mendonça-Santos-Brefin, M. L.; Montanarella, L.; Muniz-Ugarte, O.; Schad, P.; Vara-Rodríguez, M. I. and Vargas, R. 2014. Atlas de suelos de América Latina y el Caribe. Comisión Europea-Oficina de Publicaciones de la Unión Europea, L-2995 Luxembourg. 176 p.
Navarro, C. R. M.; Campo, G. A. D. y Cortina, I. S. J. 2006. Factores que afectan al éxito de una repoblación y su relación con la calidad de la planta. En: Cortina J, Peñuelas J, Puertolas J, Savé R, Vilagrosa A (coords.) Calidad de planta forestal para la restauración en ambientes mediterráneos: estado actual de conocimientos. Organimo Autómo Parques Nacionales, Ministerio de Medio Ambiente, Madrid, España. 31-46 pp.
Qin, A.; Liu, B.; Guo, Q.; Bussmann, R. W.; Ma, F.; Jian, Z.; Xu, G. and Pei, S. 2017. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch, an extremely endangered conifer from southwestern China. Global Ecology and Conservation. 10(1):139-146.
SEMARNAT. 2015. Secretaría de Medio Ambiente y Recursos Naturales. Informe de la situación del medio ambiente en México. Compendio de estadísticas ambientales. Indicadores clave, de desempeño ambiental y de crecimiento verde. 498 p. https://apps1.semarnat.gob.mx:8443/dgeia/informe15/tema/pdf/Informe15-completo.pdf.
Villar, M. R.; Ruiz, R. J.; Quero, J. L.; Poorter, H.; Valladares, R. F. y Marañón, T. 2008. Tasas de crecimiento en especies leñosas: aspectos funcionales e implicaciones ecológicas. En: Valladares F Ed. Ecología del bosque mediterráneo en un mundo cambiante. Organismo Autónomo de Parques Nacionales. Madrid, España. 193-230 pp.