elocation-id: elocation-id: e4327
La rehabilitación de suelos degradados en zonas semiáridas exige especies leñosas con alta tolerancia al estrés hídrico y potencial de provisión de servicios ecosistémicos. Prunus serotina ha sido caracterizada como tolerante al estrés hídrico y de bajas exigencias edáficas, además de presentar rápido crecimiento; sin embargo, existe escasa evidencia reciente sobre su desempeño inicial en condiciones semiáridas de México. El objetivo fue evaluar la adaptación temprana de P. serotina en campo y estimar su idoneidad ambiental regional. El estudio se realizó en 2021–2022 en Nopaltepec, Estado de México, dentro de la microcuenca del río Tula. Se determinó la supervivencia en campo a través de muestreos periódicos y se modeló la idoneidad ambiental regional mediante el algoritmo MaxEnt utilizando variables bioclimáticas y edáficas. El crecimiento vegetativo se evaluó midiendo la altura total, el diámetro basal y la cobertura de copa. Los resultados del modelo de distribución potencial indicaron una adecuada idoneidad ambiental en la región del Altiplano Central mexicano, condicionada principalmente por la estacionalidad de la precipitación y la profundidad del suelo. La supervivencia acumulada al término del periodo de evaluación fue de 74%, nivel considerado aceptable dadas las condiciones limitantes del sitio. El crecimiento en altura y diámetro basal mostró una marcada estacionalidad, con incrementos significativos durante la época de lluvias y un periodo de latencia invernal coincidente con bajas temperaturas. El análisis de riesgo mediante modelos de Cox demostró que las variables morfológicas iniciales, especialmente el diámetro del tallo, fueron determinantes en la probabilidad de supervivencia. Se concluye que P. serotina posee un alto potencial de adaptación temprana y establecimiento en suelos degradados semiáridos, lo que la convierte en una especie idónea para programas de restauración ecológica y agroforestería social en el Altiplano mexicano.
Prunus serotina, idoneidad de hábitat, riesgo de mortalidad, supervivencia.
La crisis ambiental global, intensificada por la urbanización, la expansión agrícola, los cambios en el uso de suelo y la introducción de especies exóticas invasoras, ha acelerado los procesos de degradación natural y comprometido la resiliencia de los ecosistemas. Este deterioro se manifiesta en una crisis socioecológica con efectos locales y globales, reflejada en la pérdida de biodiversidad, la reducción de servicios ecosistémicos y la disminución de la productividad de los sistemas naturales.
En México, la pérdida de cobertura natural ha sido particularmente significativa. En 1976, alrededor del 80% del territorio estaba cubierto por vegetación natural, mientras que para 2011 esta superficie se había reducido a 71.7% (140 millones de hectáreas), mientras que las áreas destinadas a agricultura, urbanización y otros usos antropogénicos aumentaron a 28% (55 millones de hectáreas). Esta tendencia se relaciona con políticas inadecuadas de uso del suelo y una planificación territorial deficiente, que han acelerado procesos de degradación con efectos potencialmente irreversibles.
La degradación del suelo constituye uno de los problemas ambientales más severos. Factores como el sobrepastoreo, la presión demográfica, la pobreza y el manejo inadecuado de la tierra aceleran la pérdida de su productividad biológica. En 2002, aproximadamente el 44.9% del territorio mexicano ya presentaba signos de degradación. Frente a esta situación, se han impulsado alternativas de producción orientadas a la rehabilitación de suelos y ecosistemas, entre las que destaca la agroforestería, que integra especies leñosas perennes con cultivos o ganado, favoreciendo la recuperación ecológica y la producción sustentable.
La selección de especies apropiadas para rehabilitación es crucial, ya que deben tolerar las condiciones locales y aportar beneficios ecológicos y productivos. Su incorporación puede evaluarse mediante Modelos de Distribución de Especies (SDM), como MaxEnt, que predicen la distribución potencial a partir de datos de presencia y variables ambientales. Estas herramientas, combinadas con análisis de supervivencia y modelos de riesgos proporcionales de Cox, permiten estimar la probabilidad de persistencia y el riesgo de mortalidad de especies bajo condiciones específicas.
En este marco, Prunus serotina (P. serotina), conocida comúnmente como capulín, se perfila como una especie con alto potencial para la rehabilitación de suelos en regiones templadas semiáridas de México, gracias a su valor alimenticio y cultural, su tolerancia a sequías y bajas temperaturas, así como a su capacidad de crecer en suelos pobres. El objetivo de este estudio fue evaluar la adaptabilidad de P. serotina en un área degradada de clima templado semiárido en la microcuenca Tula, México, mediante la integración de modelos de distribución de especies y análisis de supervivencia y crecimiento en campo, a fin de proponer su incorporación en arreglos agroforestales orientados a la rehabilitación de suelos degradados en regiones templadas semiáridas de México.
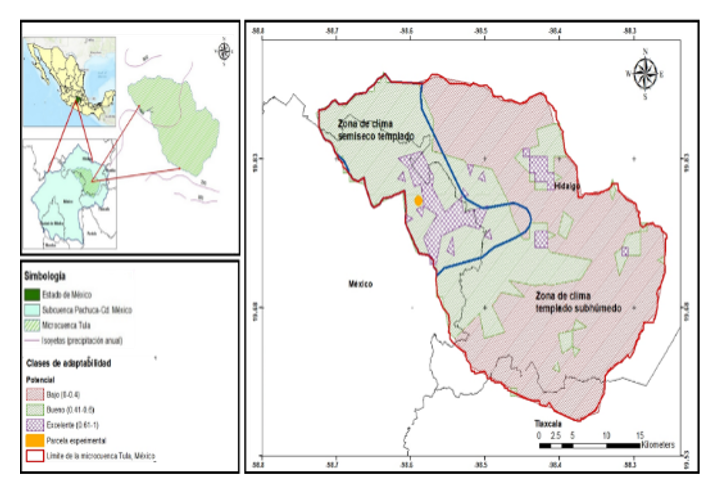
El estudio se llevó a cabo en la microcuenca de Tula, localizada entre los estados de México, Hidalgo y Tlaxcala, con una superficie aproximada de 1 037.66 km² y altitudes que oscilan entre 2 333 y 3 223 m. La precipitación media anual varía entre 500-700 mm y la temperatura entre 13-15 °C. La vegetación predominante corresponde a bosques de táscate, pino y encino y matorral crasicaule. Se distinguen dos tipos de clima: semiseco templado con lluvias en verano (28% de la superficie), con suelos Phaeozem háplico y Cambisol eutrófico; y templado subhúmedo (72%), con suelos Phaeozem háplico y Vertisol pélico.
El sitio experimental se estableció en una superficie de 1 559 m², localizada en el ejido de la comunidad de San Felipe Teotitlán, municipio de Nopaltepec, dentro de la zona semiseca de la microcuenca. El clima presenta una temperatura media anual entre 15 y 16.9 °C. La precipitación total anual varía entre 558.6 y 600 mm; la zona depende de la agricultura de temporal, y muestra procesos de degradación edáfica asociados al sobrepastoreo, tala y cambio de uso de suelo.
El experimento de campo se implementó bajo un arreglo agroforestal de árboles en lindero, con un distanciamiento de 5 m entre individuos (Figura 1). La orientación de las líneas se determinó siguiendo la dirección de la pendiente con el fin de reducir el escurrimiento superficial y la pérdida de sedimentos, además de favorecer la creación de microclimas locales. Se excavaron cepas de 30 × 30 × 40 cm, donde se trasplantaron 41 plántulas en julio de 2021.
El seguimiento se realizó durante 12 meses (julio 2021-junio 2022). En cada evaluación mensual se registró la supervivencia (1= viva, 0= muerta), la altura total (cm) con flexómetro y el diámetro del tallo (cm) con vernier digital. Cada individuo (n= 41) fue considerado como unidad de análisis.
El modelo de distribución de P. serotina se construyó a partir de registros de presencia obtenidos de GBIF (2022) y de conjunto de 19 variables bioclimáticas extraídas de la plataforma WorldClim v.2 (formatos raster con resolución espacial de 30 arcosegundos, ~1 km²). Las variables incluyeron: temperatura media anual (BIO1), rango medio diurno (BIO2), isotermalidad (BIO3), estacionalidad de la temperatura (BIO4), temperatura máxima del mes más caliente (BIO5), temperatura mínima del mes más frio (BIO6), rango anual de la temperatura (BIO7), temperatura media del trimestre más húmedo (BIO8), temperatura media del trimestre más seco (BIO9), temperatura media del trimestre más cálido (BIO10), temperatura media del trimestre más frio (BIO11), precipitación anual (BIO12), precipitación del mes más húmedo (BIO13), precipitación del mes más seco (BIO14), estacionalidad de la precipitación (BIO15), precipitación del trimestre más húmedo (BIO16), precipitación del trimestre más seco (BIO17), precipitación del trimestre más cálido (BIO18) y precipitación del trimestre más frio (BIO19). Además, se utilizó la variable elevación, obtenida de la misma plataforma, con la misma resolución espacial. Todas las variables fueron estandarizadas y processedas en formato raster.
La multicolinealidad se controló mediante correlación lineal (r≥ 0.8) en R. El modelado se realizó con MaxEnt v.3.4.1. La configuración del algoritmo fue: uso de la opción semilla aleatoria para que en cada repetición se incluyera un conjunto diferente de puntos de prueba; se utilizó el 70% de los datos para el entrenamiento y el 30% para las pruebas con remuestreo bootstrap, se utilizaron 100 repeticiones con formato logístico y un número máximo de interacciones igual a 1 000. La capacidad predictiva de los modelos se evaluó mediante el área bajo la curva (AUC) de la curva ROC; utilizando 100 réplicas, se utilizó la mediana de los valores de AUC para reducir la influencia de valores extremos. Los modelos con AUC> 0.9 tienen alta capacidad predictiva. La contribución de las variables al modelo se determinó mediante la prueba de Jackknife. Los resultados se exportaron a ArcGIS, donde se generaron un mapa continuo de idoneidad de hábitat y un mapa binario de presencia-ausencia. Se establecieron umbrales de aptitud donde valores superiores a 0.6 indican un alto potencial de plantación, valores entre 0.4 y 0.6 indican un buen potencial de plantación y valores inferiores a 0.4 indican un bajo potencial de plantación.
La supervivencia, el crecimiento y los riesgos de mortalidad de P. serotina se evaluaron en función de las variables morfológicas medidas en campo.
Estimación de la supervivencia. La tasa de supervivencia se estimó con el estimador de Kaplan-Meier, considerando observaciones completas y censuradas. La supervivencia se clasificó en cuatro rangos: excelente (≥90%), aceptable (70-89%), marginal (50-69%) y no acceptable (<49%).
Estimación del riesgo de muerte. El riesgo de muerte se evaluó mediante la regresión de riesgos proporcionales de Cox para estimar el efecto del lugar de plantación y del sistema de producción sobre las variables morfológicas de la planta como covariables. El análisis se realizó mediante el software SPSS v25.
Estimación del crecimiento vegetativo. La tasa de crecimiento en altura (HGR, cm mes-1) y la tasa de crecimiento en diámetro (DGR, mm mes-1) se calcularon utilizando ecuaciones organizadas en intervalos trimestrales correspondientes a las estaciones del año.
En la microcuenca de Tula se identificaron ocho registros de presencia de P. serotina; sin embargo, en áreas circundantes se detectaron 156 registros adicionales. Dado que la microcuenca forma parte de la cuenca Pachuca-Ciudad de México, la modelación se realizó para toda esta área (164 registros en total), y posteriormente los resultados se delimitaron a la microcuenca de Tula mediante ArcGIS.
El modelo presentó alta capacidad predictiva, con valores de AUC= 0.905 (entrenamiento) y 0.916 (prueba). Tras el análisis de correlación y el cálculo del factor de inflación de varianza (VIF), se seleccionaron seis variables no colineales para el modelado en MaxEnt: BIO1, BIO2, BIO6, BIO7, BIO12 y BIO15. La prueba Jackknife mostró que tres variables explicaron el 74.9% de la contribución total del modelo: BIO1 (42.3%); BIO12 (16.9%) y BIO7 (15.7%). Las restantes aportaron el 25.1%: BIO15 (9.3%), BIO6 (8.1%) y BIO2 (7.6%).
La Figura 2 muestra la distribución potencial de P. serotina en la microcuenca. Los resultados indican diferencias claras entre zonas climáticas: a) área semiseca templada: predominó la categoría buena (0.41-0.6) con 78.9% de la superficie, seguida de la categoría alta (0.61-1) con 17.1% y la categoría baja (0-0.4) con 4%; y b) área templado subhúmedo: predominó la categoría baja (0-0.4) con 73.5%, seguida de la categoría buena (0.41-0.6) con 25.2% y la categoría alta (0.61–1) con 1.3%.
De los 41 individuos de P. serotina establecidos en la parcela experimental, se obtuvo una tasa de supervivencia del 68.3% al término de los 12 meses de evaluación (julio 2021-junio 2022). Durante los primeros cuatro meses de evaluación se mantuvo el 100% de supervivencia, posteriormente esta comenzó a disminuir hasta alcanzar el porcentaje mencionado. Para los individuos que no murieron (censurados), el riesgo acumulado de supervivencia aumentó del mes 4 (octubre) al mes 7 (enero) en un 15% y concluyó en un 22% en el mes 12 (junio).
El modelo de Cox fue definitivo y significativo (Chi²= 19.626, p = 0.003), confirmando covariables influyentes en la supervivencia. El diámetro final mostró un efecto positivo: cada aumento de 1 cm reduce totalmente el riesgo de muerte en P. serotina, controlando otras variables (Cuadro 1).
Respecto a las tasas medios de crecimiento durante el ciclo de verano, P. serotina presentó tasas de crecimiento positivas en altura y diámetro, con 1.033 cm mes-1 y 0.201 cm mes-1. Durante el segundo período, que corresponde al ciclo otoño, la especie mostró tasas de crecimiento positivas en altura y diámetro del tallo, con 0.134 cm mes-1 y 0.176 cm mes-1, respectivamente. En el ciclo de invierno, que corresponde al tercer periodo, la especie mostró tasas de crecimiento positivas para la altura y el diámetro del tallo, con 0.201 cm mes-1 y 0.135 cm mes-1, respectivamente. Durante el cuarto período, que corresponde al ciclo primavera, la especie no mostró cambios en las tasas de crecimiento tanto en la altura (0 cm mes-1) como en el diámetro del tallo (-0.007 cm mes-1). La especie P. serotina mostró la mayor reducción en altura y diámetro en este período con respecto a los tres ciclos estacionales anteriores.
El modelo de distribución señaló que las zonas semisecas de la cuenca presentaron el mayorax potencial de plantación, lo cual concuerda con la tasa de supervivencia obtenida en el sitio experimental (68.3%), considerada marginalmente aceptable. Estos sitios se caracterizan por temperaturas medias de 14 °C, precipitaciones anuales de 500-600 mm y suelos Phaeozem háplico y Cambisol eutrófico, condiciones que se correlacionan con los hábitats naturales de la especie. El potencial de plantación estuvo fuertemente asociado a variables climáticas como BIO1 y BIO12 y a la estacionalidad entre ellas, confirmando que los regímenes de precipitación influyen de manera de forma significativa en la supervivencia.
La tasa de supervivencia observada coincide con lo destacado en la literatura sobre reforestación respecto a que la etapa de establecimiento es crítica y atribuyen la supervivencia a factores abióticos, bióticos y antrópicos. Durante los primeros tres meses, P. serotina mostró un crecimiento positivo en altura y diámetro, asociado a la temporada de lluvias, en concordancia con investigaciones que registraron un mayor crecimiento en condiciones de humedad. En otoño e invierno, el crecimiento disminuyó debido a bajas temperaturas y reducción de humedad, lo que provocó latencia fisiológica y acumulación de reservas en tallos y raíces. Al finalizar el invierno se detuvo el crecimiento, pero con la llegada de la primavera y el aumento de temperatura y precipitación, se reactivó la brotación, patrón similar al reportado en otras especies forestales.
La disminución en la tasa de crecimiento fue más evidente en altura que en diámetro, debido a que en otoño e invierno las bajas temperaturas (12-13 °C, mínimas 5-6 °C) y la menor humedad ambiental favorecieron la senescencia foliar y la latencia fisiológica, asociadas a la reducción de auxinas, giberelinas y citoquininas, y al aumento de ácido abscísico. Como respuesta adaptativa al estrés hídrico y a la escasez de nutrientes, se observó acumulación de reservas en tallos y raíces por translocación de nutrientes. Al final del invierno no se registró crecimiento, lo cual se explica porque las especies caducifolias concentran energía en romper la dormancia e iniciar la brotación de yemas. Con la llegada de la primavera, el aumento de temperatura (17 °C) y precipitación (>50 mm) favoreció la reactivación de las hormonas de crecimiento y el transporte de fotosintatos, generando condiciones opuestas a las invernales. Estudios previos en regiones similares identificaron igualmente una fase mínima de crecimiento entre octubre y mayo, seguida de un incremento con el inicio de la temporada de lluvias.
Los individuos con daños antrópicos mostraron reducciones en crecimiento, menos marcadas en el diámetro, debido a que tallos y raíces almacenan carbohidratos que sostienen el desarrollo bajo estrés. Finalmente, el análisis de riesgos de Cox confirmó que el diámetro final reduce de manera significativa el riesgo de mortalidad, lo que respalda la teoría de que algunas especies destinan más recursos al incremento diamétrico en etapas tempranas como estrategia adaptativa a las condiciones limitantes del sitio.
Este estudio demuestra que la integración de modelos de distribución de especies, junto con el análisis de supervivencia, crecimiento vegetativo y riesgo de mortalidad, constituye una herramienta eficaz para evaluar la adaptabilidad de especies multipropósito en contextos de rehabilitación de suelos degradados. La especie P. serotina mostró un potencial adecuado de plantación en las condiciones edafoclimáticas de la microcuenca de Tula, según lo mostrado por el modelo de distribución.
La tasa de supervivencia en campo fue marginalmente aceptable, lo que resalta la necesidad de considerar factores estacionales en su manejo. El crecimiento vegetativo estuvo estrechamente ligado a la estacionalidad climática: durante la temporada de lluvias se registró crecimiento positivo en altura y diámetro, mientras que en otoño e invierno las bajas temperaturas y la disminución de humedad promovieron senescencia foliar y latencia fisiológica, con acumulación de reservas en tallos y raíces. En primavera, la ausencia de crecimiento observable se asoció con el uso de esas reservas para romper la dormancia e iniciar el rebrote.
En cuanto al riesgo de mortalidad, las variables morfológicas fueron determinantes. Destaca que el diámetro final del tallo redujo en un 100% el riesgo de muerte por cada centímetro de incremento, siempre que las demás variables permanecieran constantes, lo que confirma su relevancia como indicador de adaptación temprana al sitio.
Davies, J.; Poulsen, L.; Schulte-Herbrüggen, B.; Mackinnon, K.; Crawhall, N.; Henwood, W. D.; Dudley, N.; Smith, J. and Gudka, M. 2012. Conserving dryland biodiversity. International union for conservation of nature, united nations environment programme-world conservation monitoring programme y united nations convention to combat desertification. Gland, Suiza. 84 p. https://portals.iucn.org/library/sites/library/files/documents/2012-050.pdf.
Gardi, C.; Angelini, M. E.; Barceló, S.; Comerma, J. A.; Cruz-Gaistardo, C.; Encina-Rojas, A.; Jones, A.; Krasilnikov, P.; Mendonça-Santos-Brefin, M. L.; Montanarella, L.; Muniz-Ugarte, O.; Schad, P.; Vara-Rodríguez, M. I. and Vargas, R. 2014. Atlas de suelos de América Latina y el Caribe. Comisión Europea-Oficina de Publicaciones de la Unión Europea, L-2995 Luxembourg. 176 p.
Navarro, C. R. M.; Campo, G. A. D. y Cortina, I. S. J. 2006. Factores que afectan al éxito de una repoblación y su relación con la calidad de la planta. En: Cortina J, Peñuelas J, Puertolas J, Savé R, Vilagrosa A (coords.) Calidad de planta forestal para la restauración en ambientes mediterráneos: estado actual de conocimientos. Organimo Autómo Parques Nacionales, Ministerio de Medio Ambiente, Madrid, España. 31-46 pp.
Qin, A.; Liu, B.; Guo, Q.; Bussmann, R. W.; Ma, F.; Jian, Z.; Xu, G. and Pei, S. 2017. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch, an extremely endangered conifer from southwestern China. Global Ecology and Conservation. 10(1):139-146.
SEMARNAT. 2015. Secretaría de Medio Ambiente y Recursos Naturales. Informe de la situación del medio ambiente en México. Compendio de estadísticas ambientales. Indicadores clave, de desempeño ambiental y de crecimiento verde. 498 p. https://apps1.semarnat.gob.mx:8443/dgeia/informe15/tema/pdf/Informe15-completo.pdf.
Villar, M. R.; Ruiz, R. J.; Quero, J. L.; Poorter, H.; Valladares, R. F. y Marañón, T. 2008. Tasas de crecimiento en especies leñosas: aspectos funcionales e implicaciones ecológicas. En: Valladares F Ed. Ecología del bosque mediterráneo en un mundo cambiante. Organismo Autónomo de Parques Nacionales. Madrid, España. 193-230 pp.