elocation-id: e4239
La macroalga Sargassum spp. en los últimos años se ha convertido en un problema económico y ambiental en varias regiones costeras. Sin embargo, debido a la gran variedad de compuestos bioactivos que posee, se ha propuesto como alternativa sostenible en la agricultura, ya que puede mejorar la germinación de semillas, la absorción de nutrientes, la fotosíntesis y mitigar el estrés biótico y abiótico. El objetivo de este estudio fue evaluar el efecto del cebado de semillas de tomate con extractos acuosos de Sargassum spp. sobre la germinación, vigor, biomasa, pigmentos fotosintéticos y algunos indicadores del sistema antioxidante de las plántulas. Las concentraciones de los extractos fueron: 0.5, 1.5, 2.5 y 3.5% y un control con agua destilada. Los resultados indican que los extractos mejoraron la germinación, el vigor, compuestos fenólicos, flavonoides, capacidad antioxidante y pigmentos fotosintéticos. No se observaron mejoras significativas en la biomasa. Estos resultados sugieren que la aplicación de extractos de Sargassum spp. puede ser una alternativa ecológica para mejorar los parámetros de germinación y estimular la síntesis de metabolitos secundarios en plántulas de tomate.
Solanum lycopersicum L., bioestimulantes, cebado de semillas
La agricultura moderna enfrenta el reto de incrementar la productividad y calidad de los cultivos bajo criterios de sostenibilidad y seguridad alimentaria (FAO, 2025). La creciente demanda de alimentos impulsa estrategias que optimicen el rendimiento agrícola reduciendo el impacto ambiental. En este contexto, los bioestimulantes naturales destacan por su capacidad para mejorar la producción y calidad, disminuyendo el uso de agroquímicos y mitigando el estrés biótico y abiótico (Durán-Hernández et al., 2022). Entre ellos, los derivados de algas marinas, como Sargassum spp., son prometedores por su riqueza de compuestos bioactivos. Estas macroalgas contienen polisacáridos, compuestos fenólicos, carotenoides, fitohormonas y aminoácidos, que mejoran la germinación, el crecimiento y calidad, además de conferir tolerancia al estrés salino, hídrico, térmico y por patógenos (Senthilkumar et al., 2024).
Aunque su proliferación masiva genera problemas ambientales y económicos en el Caribe Mexicano. Su alto contenido de metabolitos secundarios la convierte en una fuente valiosa de bioestimulantes agrícolas (Adderley et al., 2023). Estudios previos demuestran su potencial para mejorar el crecimiento y la calidad nutracéutica de distintos cultivos (Rivera-Solís et al., 2021).
Por su parte, el tomate (Solanum lycopersicum L.) es una de las hortalizas más cultivadas y consumidas mundialmente, apreciado por su valor nutricional y antioxidante (Collins et al., 2022). Para asegurar la productividad de este cultivo, se requieren semillas de alta calidad que garanticen una germinación uniforme, etapa clave para el rendimiento del cultivo (Ruiz-Ramirez et al., 2021; Reed et al., 2022). La germinación, transición de latencia a crecimiento activo, puede optimizarse mediante tratamientos pregerminativos como el cebado, que mejora la tasa de germinación, vigor y tolerancia al estrés (Wang and Shi, 2024; Abinandan et al., 2025). Dado el potencial bioestimulante del Sargassum spp., este estudio evaluó el efecto de extractos de Sargassum spp. sobre la germinación, pigmentos fotosintéticos y actividad antioxidante en plántulas de tomate.
Se utilizó alga parda Sargassum spp., previamente deshidratada y pulverizada, proporcionada por la Universidad Autónoma Agraria Antonio Narro (UAAAN). El extracto líquido se preparó según la metodología de Sariñana-Aldaco et al. (2021), donde se detalla su caracterización bioquímica. Se elaboró una solución madre (100%) en proporción 1:20 (m:v, alga:agua destilada), hervida durante 1 h, prensada y filtrada. A partir de esta solución concentrada se obtuvieron las diluciones requeridas (0.5, 1.5, 2.5 y 3.5%) con agua bidestilada.
Se utilizaron semillas de tomate tipo saladette variedad ‘Río Grande’, desinfectadas con etanol al 75% durante 5 min y lavadas dos veces con agua destilada. Las semillas se sumergieron durante 24 h en las concentraciones del extracto de Sargassum spp. (0.5, 1.5, 2.5 y 3.5%), incluyendo un control con agua destilada. Posteriormente, se colocaron en papel de filtro #1 dentro de cajas Petri estériles (90 mm), con 10 semillas por caja y 20 cajas por tratamiento. El papel de filtro se humedeció con agua destilada estéril. El diseño experimental fue completamente al azar con cinco tratamientos y 20 repeticiones. Las cajas Petri se incubaron en una cámara de germinación (Achieva, Lab-Tech Inc, Modelo Núm. A-3920), bajo ciclo de día/noche de 12 h, a 25 ±2 °C y 60% de humedad relativa (Li et al., 2019). Tras 14 días, se evaluaron porcentaje de germinación, la biomasa fresca y el porcentaje de vigor.
El porcentaje de germinación (G) y de vigor (V) fueron calculados mediante las siguientes fórmulas:
Donde: n= número de semillas germinadas en el día 14; N= número total de semillas.
Donde: Pn= plántulas normales; N= número total de semillas.
El contenido de clorofila se determinó según el método de Lichtenthaler y Wellburn (1983), utilizando etanol al 95% como disolvente. Las lecturas de absorbancia se realizaron a 665, 649 y 470 nm en un espectrofotómetro UV-visible (Jenway 7305).
Las concentraciones de clorofila se calcularon mediante las siguientes ecuaciones:
Se homogeneizaron 2 g de muestra fresca con 10 ml de etanol al 80%, mediante agitación orbital constante (70 rpm) durante 24 h a 70 rpm y 5 °C. Posteriormente, los extractos fueron centrifugados a 3 000 rpm durante 5 min, el sobrenadante se utilizó para los análisis posteriores.
El contenido de compuestos fenólicos se determinó mediante una adaptación del método Folin-Ciocalteu (Singleton et al., 1999). Se mezclaron 50 µl de extracto etanólico con 3 ml de agua destilada y 250 µl de reactivo Folin-Ciocalteu (1 N). Tras 3 min de reposo, se añadieron 750 µl de Na2CO3 y 950 µl de agua destilada, incubando 2 h. La absorbancia se midió a 765 nm en un espectrofotómetro UV-visible (Jenway 7305), expresando los resultados como mg equivalentes de ácido gálico (AG) por cada 100 g de peso fresco (PF).
El contenido total de flavonoides se cuantificó mediante el método colorimétrico descrito por Buendía-García et al. (2021). Se mezclaron 250 µl de extracto con 1.25 ml de agua destilada y 75 µl de NaNO2 (5%), reposando 5 min. Luego se añadieron 150 µl de AlCl3, 500 µl de NaOH (1M) y 275 µl de agua destilada. La absorbancia se midió a 510 nm, expresando los resultados se expresaron en mg equivalentes de quercetina (QE) por cada 100 g PF.
La capacidad antioxidante se evaluó mediante el método DPPH+ (Brand-Williams et al., 1995), utilizando una solución de DPPH+ en etanol (0.025 mg ml-1). Se mezclaron 50 µl con 1 950 µl de DPPH+, incubando 30 min y se midió la absorbancia a 517 nm. Los resultados se expresaron en miliequivalentes (Meq) de Trolox por cada 100 g de peso PF.
La actividad de la catalasa (CAT, EC 1.11.1.6) se determinó según el método de Aebi (1974). Las lecturas se realizaron a temperatura ambiente en un espectrofotómetro UV-Visible (Jenway 7305), monitoreando la disminución de la absorbancia a 240 nm por descomposición del H2O2. La actividad enzimática se calculó empleando el coeficiente de extinción molar (ε240 = 43.6 M cm-1) y la concentración de proteínas, determinada por el método de Bradford (1976). Los resultados se expresaron como U mg-1 proteína, definiendo una unidad (U) como la cantidad de enzima que provoca un cambio de 0.001 de absorbancia por minuto en las condiciones del ensayo.
La normalidad de los datos se verificó mediante la prueba de Kolmogorov-Smirnov. Los datos expresados en porcentaje se transformaron mediante funciones de arco seno o raíz cuadrada, según su distribución. El análisis de varianza y la comparación de medias mediante la prueba de Tukey (p ≤ 0.05) se realizaron con el paquete estadístico Statistical Analysis System (SAS), versión 9.3.
La germinación y el vigor son etapas críticas en el desarrollo vegetal, derminantes para el establecimiento y el rendimiento del cultivo (Papoui et al., 2025). En este estudio, el tratamiento con el extracto líquido de Sargassum spp. mejoró significativamente la germinación y el vigor de las semillas de tomate (Figura 1). La concentración de 3.5% incrementó la germinación en 19%, mientras que 1.5% aumentó el vigor en 13% respecto al control.
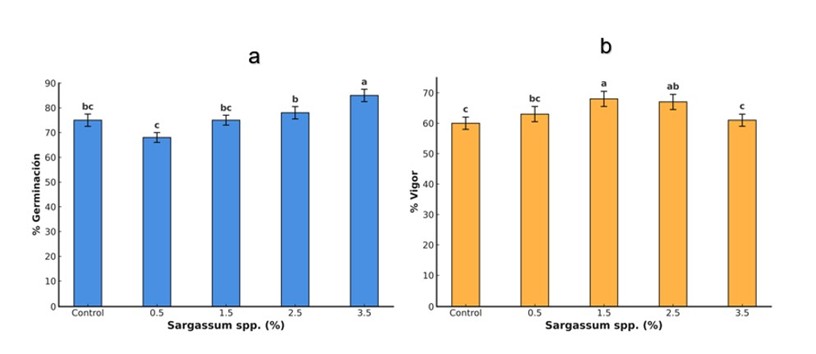
Resultados similares han sido reportados en tomate con extractos de algas pardas (Hernández-Herrera et al., 2023) y en Abelmoschus esculentus con extractos de Ecklonia maxima (Makhaye et al., 2021). El efecto positivo se atribuye a la presencia de fitohormonas, polisacáridos y compuestos antioxidantes que regulan procesos clave durante la germinación (Cossa et al., 2023; Margal et al., 2023). Las algas pardas, que contienen auxinas, giberelinas y citoquininas que facilitan la ruptura de la dormancia mediante la activación de enzimas hidrolíticas, como la amilasa, que convierte el almidón en azúcares simples para el desarrollo embrionario (Sariñana-Aldaco et al., 2022). Además, estas fitohormonas promueven la división y elongación celular (Zluhan-Martínez et al., 2021). No obstante, su efectividad depende de múltiples factores, como la especie de alga, tipo de extracción, condiciones ambientales, concentración y la especie vegetal receptora (Martínez-González et al., 2022). Concentraciones elevadas pueden generar efectos adversos por acumulación de compuestos que alteran el equilibrio hormonal o inducen estrés fisiológico (Castro et al., 2022).
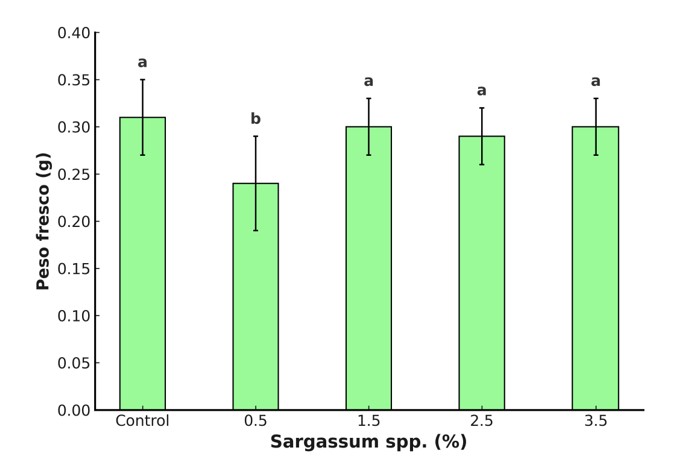
El cebado con extracto líquido de Sargassum spp. no causó efectos significativos sobre la biomasa (Figura 2). Estos resultados coinciden con estudios previos que no reportan diferencias entre distintas concentraciones utilizadas. Por ejemplo, Morales-Meléndez et al. (2023) no observaron incrementos significativos en la producción de biomasa de plantas de tomate tratadas con extractos de algas y Rivera-Solís et al. (2021) tampoco encontraron diferencias respecto al control. Esto sugiere que la composición química de los extractos y su concentración son factores clave para inducir una respuesta fisiológica positiva o evitar efectos inhibitorios.
La clorofila total y los carotenoides son indicadores clave de la capacidad fotosintética y del estado fisiológico de las plantas (Sherin et al., 2022). En este estudio, ambos pigmentos fueron significativamente afectados por los extractos de Sargassum spp. (Cuadro 1). La concentración de 3.5% redujo la clorofila total en 14.85% respecto al control. Resultados similares han sido reportados con extractos de algas marinas (Salazar-Salazar et al., 2022; Mohammed et al., 2023).
| Extracto de Sargassum spp. (%) | Clorofila total (mg g-1 PS) | Carotenoides (mg g-1 PS) |
|---|---|---|
| Control | 1.28 b | 102.68 b |
| 0.5 | 1.48 ab | 118.27 ab |
| 1.5 | 1.91 a | 137.8 a |
| 2.5 | 1.61 a | 128.44 ab |
| 3.5 | 1.09 b | 82.91 b |
Este efecto se ha atribuido a la acción de betaínas presentes en los extractos, que actúan como chaperonas moleculares estabilizando biomoléculas sensibles, como los pigmentos y evitando su degradación (Sariñana-Aldaco et al., 2025). Dichos compuestos han demostrado prevenir la pérdida de actividad fotosintética al inhibir la degradación de la clorofila (Genard et al., 1991), especialmente en dosis intermedias.
El sistema antioxidante vegetal comprende dos componentes: enzimático, considerado la primera línea de defensa, integrado por enzimas como catalasa (CAT), ascorbato peroxidasa (APX) y glutatión peroxidasa (GPX), que degradan especies reactivas de oxígeno (ROS) como H2O2 (Dumanović et al., 2021) y no enzimático, conformado por metabolitos antioxidantes como ácido ascórbico, compuestos fenólicos, glutatión y carotenoides (Hashim et al., 2020). En este estudio, el extracto de Sargassum spp. (1.5%) incrementó significativamente fenoles totales (141%), flavonoides (125%) y capacidad antioxidante (11%) respecto al control (Figura 3). Sin embargo, la actividad de CAT disminuyó en tratamientos con 1.5% y 3.5% (Figura 4). Estos resultados concuerdan con reportes sobre el efecto positivo de extractos de algas en la acumulación de compuestos bioactivos (Sariñana-Aldaco et al., 2021), aunque difieren de estudios que informan aumentos en enzimas antioxidantes en soya tratada con extractos de Ascophyllum nodosum (Repke et al., 2022).
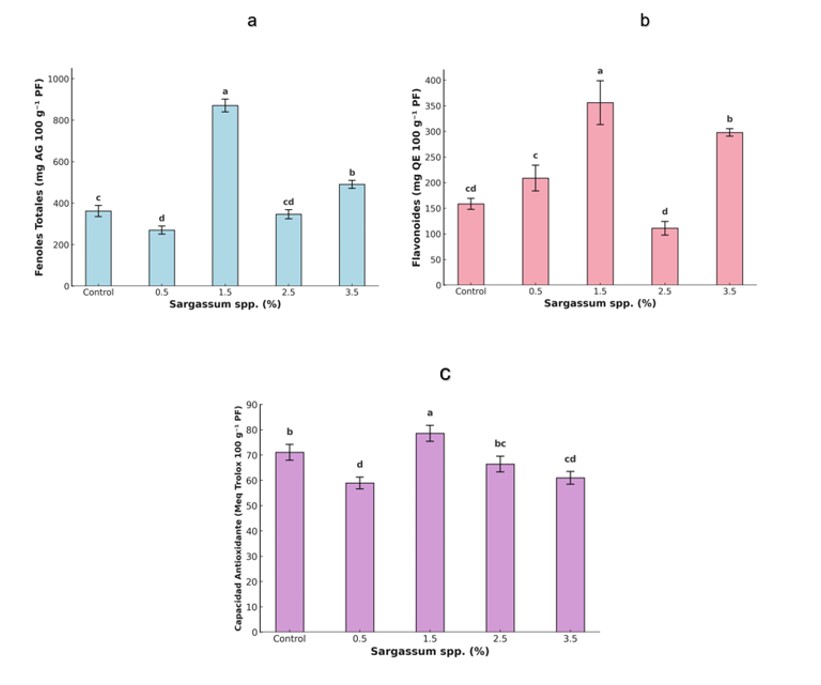
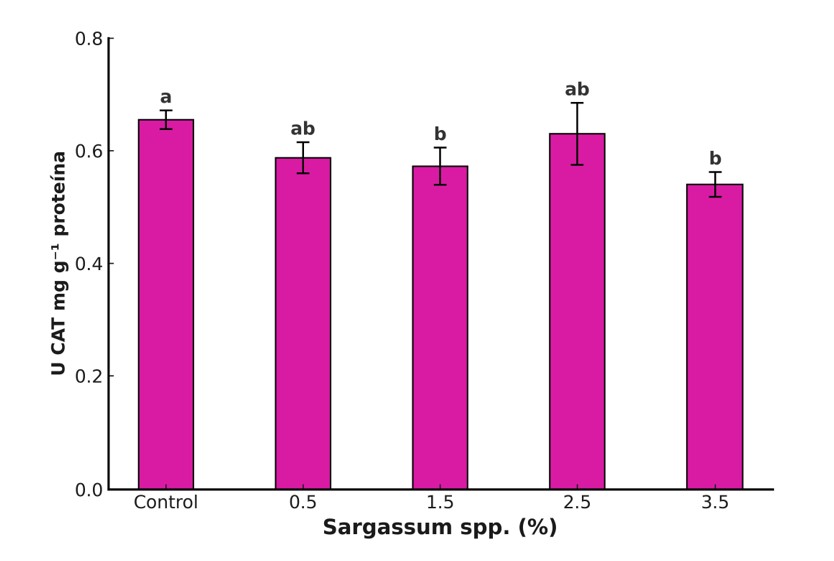
La respuesta observada puede atribuirse a metabolitos del extracto que activan cascadas de señalización y expresión génica para enzimas antioxidantes y síntesis de compuestos fenólicos (González-Morales et al., 2021). Además, algunos metabolitos ejercen acción directa en el citoplasma (Lau et al., 2025). La reducción en CAT sugiere un reequilibrio del sistema antioxidante, favoreciendo la defensa no enzimática mediante la biosíntesis de fenoles y flavonoides, lo que disminuye la necesidad de rutas enzimáticas para neutralizar ROS (Kumar et al., 2024).
El extracto de Sargassum spp. como bioestimulante natural mejoró la germinación de las semillas de tomate, el vigor de las plántulas y la biosíntesis de compuestos bioactivos, incrementando su capacidad antioxidante. El extracto de Sargassum spp. es una alternativa sostenible para favorecer la germinación, el desarrollo de plántulas, así como mayor producción de metabolitos secundarios.
Adderley, A.; Wallace, S.; Stubbs, D.; Bowen-O’Connor, C.; Ferguson, J.; Watson, C. and Gustave, W. 2023. Sargassum sp. as a biofertilizer: is it really a key towards sustainable agriculture for The Bahamas? Bulletin of the National Research Centre. 47(1):1-11. https://doi.org/10.1186/s42269-023-01087-w.
Buendía-García, A.; Lozano-Cavazos, C. J.; Rodríguez-Ortiz, J. C.; Carballo-Méndez, F. D. J.; Moreno-Reséndez, A.; Sariñana-Navarrete, M. D. A. y Preciado-Rangel, P. 2021. La biofortificación con selenio mejora la calidad nutracéutica y la capacidad antioxidante de la lechuga (Lactuca sativa L.). Ecosistemas y Recursos Agropecuarios. 8(3):1-8. https://doi.org/10.19136/era.a8n3.3104.
Hernández-Herrera, R. M.; González-González, M. F.; Velasco-Ramírez, A. P.; Velasco-Ramírez, S. F.; Santacruz-Ruvalcaba, F. and Zamora-Natera, J. F. 2023. Seaweed extract components are correlated with the seeds germination and growth of tomato seedlings. Seeds. 2(4):436-448. https://doi.org/10.3390/seeds2040033.
Mohammed, S.; El-Sheekh, M. M.; Aly, S. H.; Al-Harbi, M.; Elkelish, A. and Nagah, A. 2023. Inductive role of the brown alga Sargassum polycystum on growth and biosynthesis of imperative metabolites and antioxidants of two crop plants. Frontiers in Plant Science. 14(11):1-12. https://doi.org/10.3389/fpls.2023.1136325.
Morales-Meléndez, R.; Betancourt-Galindo, R.; Juárez-Maldonado, A.; Hernández-Pérez, A.; González-Fuentes, J. A.; Puente-Urbina, B. y Méndez-López, A. 2023. Aplicación de extractos de algas, NP’SZnO y microorganismos sobre la biomasa vegetal en tomate. Ecosistemas y Recursos Agropecuarios. 10(2):1-11. https://doi.org/10.19136/era.a10n2.3206.
Rivera-Solís, L. L.; Rodríguez-Jasso, R. M.; Flores-López, M. L.; Robledo-Olivo, A.; Sandoval-Rangel, A.; Sariñana-Aldaco, O. y González-Morales, S. 2021. Extractos de Sargassum spp. como inductores de tolerancia a Fusarium oxysporum en plántulas de tomate. Ecosistemas y Recursos Agropecuarios. 8(1):1-8. https://doi.org/10.19136/era.a8n1.2826.
Sariñana-Aldaco, O.; Benavides-Mendoza, A.; Juarez-Maldonado, A.; Robledo-Olivo, A.; Rodríguez-Jasso, R. M.; Preciado-Rangel, P. y Gonzalez-Morales, S. 2021. Efecto de extractos de Sargassum spp. en el crecimiento y antioxidantes de plántulas de tomate. Ecosistemas y Recursos Agropecuarios. 8(2):1-15. https://doi.org/10.19136/era.a8n2.2814.
Sariñana-Aldaco, O.; Rivera-Solís, L. L.; Benavides-Mendoza, A.; Robledo-Olivo, A., Rodríguez-Jasso, R. M. and González-Morales, S. 2025. Using brown algae in the plant-soil system: a sustainable approach to improving the yield and quality of agricultural crops. Horticulturae. 11(1):1-29. https://doi.org/10.3390/horticulturae11010094.
Senthilkumar, S.; Kuppusamy, S.; Palai, S.; Rajanbabu, V. and Vennila, A. 2024. Exploring Sargassum extract as a bio stimulant: advancements and benefits in soil plant systems a review. Communications in Soil Science and Plant Analysis. 55(22):3638-3648. https://doi.org/10.1080/00103624.2024.2397013.
Zluhan-Martínez, E.; López-Ruíz, B. A.; García-Gómez, M. L.; García-Ponce, B.; De la Paz Sánchez, M.; Álvarez-Buylla, E. R. and Garay-Arroyo, A. 2021. Integrative roles of phytohormones on cell proliferation, elongation and differentiation in the Arabidopsis thaliana primary root. Frontiers in Plant Science. 12(6):1-20. https://doi.org/10.3389/fpls.2021.659155.