elocation-id: elocation-id: e4017
Beans are one of the main components of traditional Mexican gastronomy. Mexico is the center of origin, domestication, and diversification of Phaseolus vulgaris and P. coccineus. These are cultivated in the Ñäñho communities of Amealco, Querétaro, Mexico; however, their characteristics are unknown. The work aimed to determine inter and intraspecific seed diversity based on morphological characteristics and chemical composition. The study was conducted in 2024 at the Faculty of Natural Sciences of the Autonomous University of Querétaro. Twenty-one accessions (eight of P. coccineus and 13 of P. vulgaris) held in safekeeping at the Germplasm Bank-UAQ, which were collected in 2021 and 2022 in San Miguel Tlaxcaltepec, San Ildefonso and Santiago Mexquititlán, communities of the municipality of Amealco, Querétaro, were analyzed. Four quantitative morphological variables and 11 qualitative variables of each accession’s seeds were measured. In addition, chemical analysis was carried out, determining ash, moisture, crude protein, ethereal extract, crude fiber and nitrogen-free extract contents. Statistically significant differences were found in size, weight and protein contents between the two species. Intraspecific diversity was identified in the two species, mainly defined by seed color and size, as well as by percentages of ethereal extract and crude fiber. The Ñäñho communities of Amealco, Querétaro, present inter- and intraspecific diversity in both species, with a higher protein content in P. vulgaris than in P. coccineus.
P. coccineus, P. vulgaris, chemical, morphology.
Beans (Phaseolus spp.) are the second most important crop in Mexico and are a relevant source of protein, starch, vitamins and fiber (Aquino-Bolaños et al., 2021). The genus Phaseolus is native to the Americas and comprises 77 species (López-Báez et al., 2018). Of the species recorded in Mexico, five have been domesticated, including P. vulgaris L. (common beans) and P. coccineus L. (ayocote beans) (Ayala-Garay et al., 2021).
P. vulgaris is considered to have been domesticated in Mesoamerica and the Andes (Castillo-Mendoza et al., 2006), whereas for P. coccineus, Mexico is considered the only center of domestication (Vargas-Vázquez et al., 2014). In the state of Querétaro, 12 bean species are found (Quiroz-Sodi et al., 2018). In the Ñäñho communities of the municipality of Amealco, Querétaro, Hernández-Sandoval et al. (2023); Hernández-Puente et al. (2025) mention the presence of P. coccineus and 18 common names of P. vulgaris.
Crop diversity is essential for their adaptation to the effects of climate change and for meeting different human needs (Vargas-Vázquez et al., 2014). In studies of populations of P. vulgaris and P. coccineus in Puebla, State of Mexico, Guerrero and Tlaxcala, based on morphoagronomic characterization, it was possible to identify interspecific and intraspecific diversity (Castillo-Mendoza et al., 2006; Vargas-Vázquez et al., 2014; Espinosa-Pérez et al., 2015). Specifically, seed-related traits enabled the grouping of 75 populations from south-central Mexico (Espinosa-Pérez et al., 2015).
Statistically significant differences were recorded between P. vulgaris and P. coccineus in the chemical composition of seeds, including the contents of proteins, sugars, phenols and secondary metabolites (Quiroz-Sodi et al., 2018; Pérez-Herrera et al., 2002; Aquino-Bolaños et al., 2021).
There is also intraspecific diversity in both species. Fernández-Valenciano and Sánchez-Chávez (2017) found significant differences in the physicochemical properties of improved varieties of P. vulgaris in fiber, protein and iron contents. In this same species, Granito et al. (2009) noted differences in nutrient content between raw and cooked samples. A study by Teniente-Martínez et al. (2006) found differences in the protein content of black (23.8%) and purple (21.9%) P. coccineus from the Northern Sierra of the state of Puebla. Nonetheless, purple beans contain higher levels of ash, fat and carbohydrates.
In 2023, bean production was 724 000 t (SIAP, 2024), the lowest reported in the last 30 years. The genetic richness of beans faces serious problems due to changes in consumption habits, industrialization, which affects the countryside and the substitution of food for fast food. The use of monocultures has reduced the use of varieties associated with native corn within the milpa. Given the above, the present study performed a morphological and chemical characterization of native seeds of P. vulgaris and P. coccineus from the Ñäñho communities of Amealco, Querétaro, to identify the inter- and intraspecific diversity that farmers conserve and select.
The morphological characterization and proximate chemical analysis were carried out in 2024 at the Faculty of Natural Sciences of the Autonomous University of Querétaro (UAQ). Seeds held in safekeeping at the Germplasm Bank-UAQ were used. Collections of P. vulgaris and P. coccineus from three Ñäñho indigenous communities, corresponding to San Miguel Tlaxcaltepec, San Ildefonso Tultepec and Santiago Mexquititlán in the municipality of Amealco de Bonfil, Querétaro, Mexico, were selected.
The most recent collections, corresponding to the years 2021 and 2022, were selected. A total of 21 accessions were characterized, of which eight belonged to the species P. coccineus (PC) and 13 to P. vulgaris (PV). The common names for the PC accessions stated in the passport data were bayocote, frijol grande and burro. In the case of PV, some common names are ojo de cabra for accession PV2, cacahuate for PV7, rojo for PV8, san franciscano for PV10 and PV11 and vaquita for PV13.
For the morphological analysis, 50 seeds per accession were used. Four quantitative morphological variables were evaluated: weight, thickness, width, and length. Likewise, 11 qualitative variables were characterized, with their different trait states, according to the technical guide for varietal description of beans (Phaseolus vulgaris L.) (SNICS, 2017) (Table 1).
Color measurements were taken with a colorimeter (Precise Color Reader, WR-10), which provides value in wavelengths L, a and b. To characterize the shape in the longitudinal and transversal sections, half of the material was used, 200 seeds for P. coccineus and 325 for P. vulgaris, with 25 seeds per accession. A digital vernier caliper (STAINLESS HARDENED.CAT.CA3930) and an analytical balance (OHAUS Item Pa31) were used for all morphometric measurements.
The chemical composition of beans was determined according to the Association of Official Analytical Chemists (AOAC, 2000). The bean samples were ground in an electric mill (IKA Works, Inc. A10 52) and then used for the following analyses.
Ash content (A). Ash is the inorganic part of food left after burning it at high temperatures. The analysis was performed by incinerating the samples in a muffle at 550-600 °C for 6 h. The percentage was calculated as the weight difference between the calcined and initial samples, multiplied by 100.
Moisture content (ME). It allowed the water content to be determined. The samples were placed in the drying oven at 105 °C for 4 h. The moisture percentage was obtained by weight difference, where the weight of the crucible with the dry sample was subtracted from the weight of the sample plus the weight of the crucible, then divided by the total weight of the sample and multiplied by 100.
Crude protein (CP) content. It determines the amount of N present in organic and inorganic samples. The Kjeldahl method was used, which consists of three steps: digestion, distillation and titration (AOAC, 2000). The protein concentration was expressed as a percentage, considering a protein factor of 6.25.
Ethereal extract (EE) content. The total ether-soluble lipid content was specified. A total of 1.5 g of each sample was weighed and placed in the base of the BUCHI fat extractor, where 60-80 ml of petroleum ether was added. The concentration was expressed as a percentage by weight differences.
Crude fiber (CF) content. The contents of cellulose, hemicellulose, and lignin present in the cell walls were determined. For each sample, a filter bag was weighed, 0.5 g was placed in it, the bag was sealed with heat, and then it was placed in a digester. The percentage of fiber was determined by weight difference.
Nitrogen-free extract (NFE) content. This determination quantifies the content of non-structural carbohydrates. This fraction is calculated by subtracting the sum of the total percentages from 100: 100 - (CP% + CF% + A% + EE% + ME%).
Statistical analysis. To identify the statistical differences between the two species, the U-Mann-Whitney test was applied, as the data did not present a normal distribution. To evaluate intraspecific diversity within each species, multivariate cluster analyses were conducted using the WardD method and Euclidean distance. Non-metric multidimensional scaling (NMDS) ordering analysis was also used to identify the main variables that allow accessions to be grouped. All statistical analyses were performed in R version 4.
Statistically significant differences were found between the two bean species across the quantitative and qualitative variables. For the traits of weight, length, thickness and width, P. coccineus was statistically larger than P. vulgaris. For the color intensity trait, which was determined through wavelengths, no statistically significant differences were found for variable a, while variables L and b showed differences (Table 2).
Differences were found in the qualitative characteristics of the seed. For the shape of the longitudinal section (SLS) in both species, five of the six trait states considered by the SNICS (2017) varietal description technical guide were found. In P. vulgaris, the predominant shape was circular with 64 seeds (33%), whereas in P. coccineus, it was rectangular with 47 seeds (45%).
Similarly, in the shape of the transversal section (STS), the five trait states considered by the technical guide were present in both species. However, there are differences between the two species; the most common shape was the flattened one, with 73 seeds (47%) corresponding to P. vulgaris; for P. coccineus, the most predominant shape was the circular one, with 38 seeds (40%).
Regarding the number of colors (NCOL), P. vulgaris presented greater diversity, with seven colors and P. coccineus showed five colors. The testa brightness and venation are higher in P. coccineus than in P. vulgaris, but for the color of the hilum crown, the same color as in the testa predominates in both (Figures 1a and 1b).

A greater diversity of colors was found in P. vulgaris than in P. coccineus, similar to that reported by Castillo-Mendoza et al. (2006) in the study of the morphological diversity of beans in northern Morelos and the east of the State of Mexico; the predominance of certain colors may depend on the region and use.
As observed in Table 2, P. coccineus has greater intraspecific diversity. In terms of weight, length, thickness, and width, this species has higher CV values than P. vulgaris. In weight, P. coccineus has a minimum of 0.25 g and a maximum of 2 g (CV: 51%); on the other hand, P. vulgaris shows values of 0.09 to 0.6 g (CV: 31%). In length, P. coccineus has values of 8.8 to 22.6 mm (CV: 20%), and P. vulgaris has values of 3.78 to 13.14 mm (CV: 12%).
Different morphological characterization studies in P. vulgaris and P. coccineus showed that some of the main traits that allow the grouping of the different populations are the characteristics related to the color, size, weight, and geographical origin of the seed (Castillo-Mendoza et al., 2006; Espinoza-Pérez et al., 2015; López-Báez et al., 2018). This may be because, during domestication, the main selection criteria are seed size, color and earliness (López-Báez et al., 2018).
In the case of the diversity of colors present in both species, it depends on the region and the uses of the seed, whether in traditional gastronomy or in cultivation (Vargas-Vázquez et al., 2012). Intraspecific morphological differences may be due to the ranges of adaptation to different environmental conditions and to biological aspects of the species, since P. vulgaris is autogamous and has lower CVs than P. coccineus, an open-pollinated, allogamous species (López-Báez et al., 2018; Ayala-Garay et al., 2021).
No statistically significant differences were found in the contents of moisture, ash, ethereal extract, crude fiber and nitrogen-free extract between P. vulgaris and P. coccineus. Nevertheless, they differed significantly in the percentage of crude protein (Table 3).
As shown in Table 3, the percentages of moisture, ash, crude protein and nitrogen-free extract have CVs equal to or less than 10 in both species. Therefore, the contents are similar across the different accessions, as indicated by the minimum and maximum values obtained.
On the contrary, in the percentages of ethereal extract and crude fiber, CVs range from 28% to 43%, so there is intraspecific diversity in both species. In the case of P. vulgaris, accession PV9 had the lowest ethereal extract content at 0.67%, and accession PV11 (san franciscano) had the highest at 2.78%.
In the case of P. coccineus, the accession with the lowest value was PC2 (white bayocote) at 0.31%, and the highest was PC6 (purple bayocote) at 2.12% (Figures 2a and 2c).
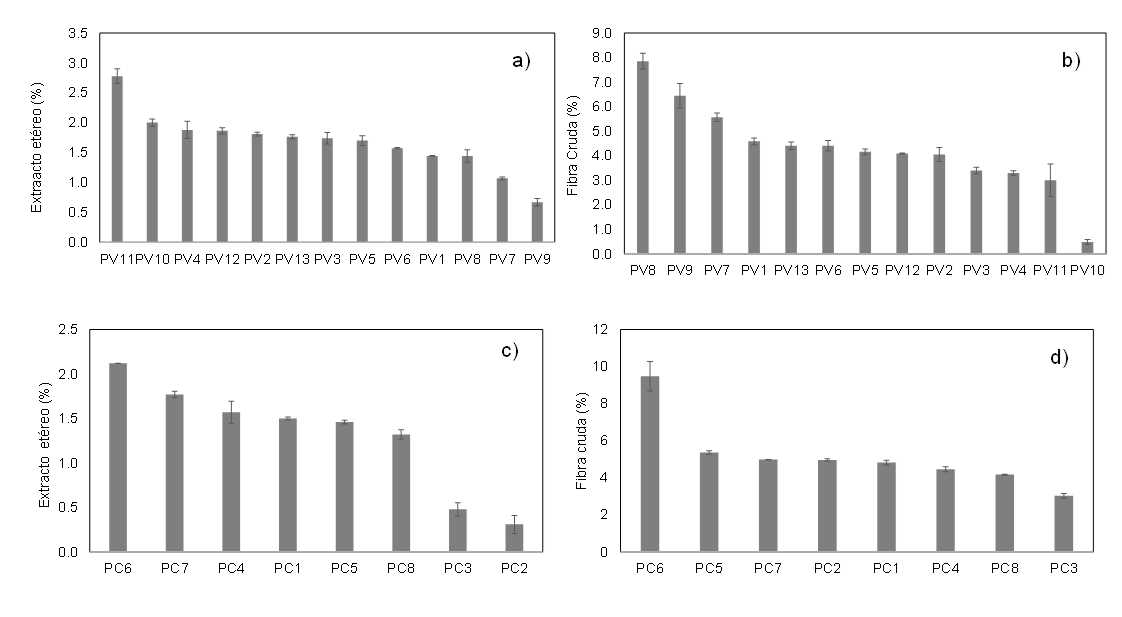
In the percentage of crude fiber, the P. vulgaris accession with the lowest value was PV10 (san franciscano), at 0.49%, which was statistically different from the content of accession PV8, which had the highest value, 7.86%. In P. coccineus, the accession with the lowest percentage was PC3 (3.03%), and the highest was PC6 (purple bayocote) (9.48%), with statistically significant differences (Figures 2b and 2d).
Aquino-Bolaños et al. (2021) and Pérez-Herrera et al. (2002) also indicate a higher percentage of crude protein content in P. vulgaris than in P. coccineus. Regarding the content of ethereal extract, Granito et al. (2009); Teniente-Martínez et al. (2006); Pliego et al. (2013) mention that it can range from 1.2 to 3.78% in P. vulgaris and P. coccineus.
On the other hand, in six improved varieties of P. vulgaris, the percentages of crude fiber can be between 5.55 and 11.35% (Fernández-Valenciano and Sánchez-Chávez, 2017). These wide intervals may be due to aspects of farmer selection and management, as well as to the availability of nutrients in the planting areas, as it affects the concentration of some of these compounds (Granito et al., 2009).
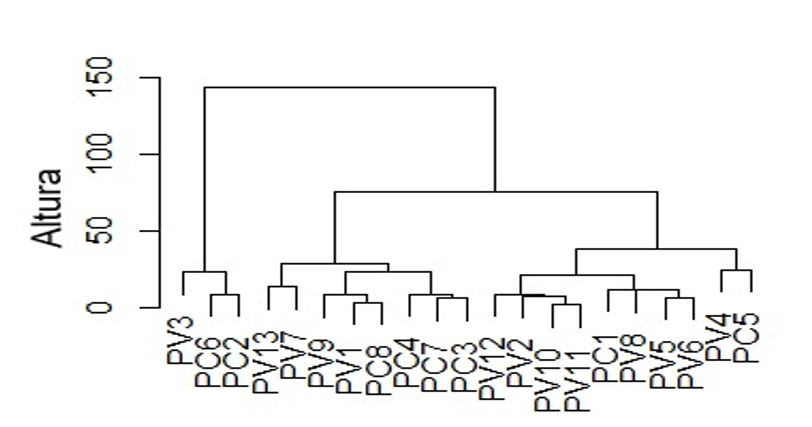
Considering quantitative morphological variables and chemical composition results, the cluster analysis of similarity between the two species yields two groups. However, the two species did not separate. Group one is made up of accessions PC6 and PC2 of P. coccineus and accession PV3 of P. vulgaris.
Group two has two subgroups, one with four accessions of P. vulgaris and four of P. coccineus, and the other with eight accessions of the first species and two of the second species (Figure 3). This is because there are morphological differences, but in chemical composition, they are similar. According to the NMDS analysis (stress: 0.09), the variables that allow the grouping of accessions are seed size, seed thickness and seed color.
This intraspecific diversity coincides with studies by Hernández-Sandoval et al. (2023); Hernández-Puente et al. (2025), who report the presence of different traditional varieties in the study communities. Espinoza-Pérez et al. (2015), also based on morphological characterization of the seed, grouped 75 populations of P. vulgaris from south-central Mexico into 13 classes.
For this same species, Vargas-Vázquez et al. (2012) grouped 200 populations from the INIFAP core collection into five classes. Vargas-Vázquez et al. (2014), in the morphoagronomic characterization of P. coccineus, grouped the populations into four classes based on their geographical origin, earliness, and seed size.
Inter- and intraspecific diversity was found between P. vulgaris and P. coccineus from the three Ñäñho communities of Amealco, Querétaro. The main traits that grouped the collections were seed color, size and weight, ethereal extract and crude fiber.
A higher protein percentage content and greater intraspecific diversity for P. vulgaris stand out. The morphological traits and chemical composition of the seeds allowed us to differentiate groups within both P. vulgaris and P. coccineus, but without forming different groups between the two.
These materials can be a source of genes for genetic improvement programs to generate highly nutritious varieties.
Aquino-Bolaños, E.; Garzón-García, A. K.; Alba-Jiménez, J. E.; Chávez-Servia, J. L.; Vera-Guzmán, A. M.; Carrillo-Rodríguez, J. C. and Santos-Basurto, M. A. 2021. Physicochemical characterization and functional potential of Phaseolus vulgaris L. and Phaseolus coccineus L. landrace green beans. Agronomy. 11(4):803. Doi: 10.3390/agronomy11040803.
Hernández-Puente, K. N.; Hernández-Sandoval, L.; González-Santos, R.; Casas, A.; Martínez, M. and Steimann, V. W. 2025. Diversity management and uses of edible plants in a Ñäñho community of Southern Querétaro, México. Journal of Ethnobiology and Ethnomedicine 21(1):18. Doi: https://pmc.ncbi.nlm.nih.gov/articles/PMC11899470/.
SNICS. 2017. Servicio Nacional de Inspección y Certificación de Semillas. Guía técnica para la descripción varietal. (Phaseolus vulgaris L). Servicio Nacional de Inspección y Certificación de Semillas (SNICS). Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA). México, DF. 36 p.