elocation-id: e3722
Damping-off is a disease that occurs in forest nurseries during the establishment stage of seedlings and is mainly caused by fungi. The objective was to carry out the isolation, morphological characterization, and pathogenicity tests of fungi in five forest nurseries in the state of Durango in 2018. Fungal isolates were obtained from seedlings of Pinus engelmannii, P. durangensis, P cooperi, and P. arizonica approximately 45 days old, which were selected based on the presence of symptoms characteristic of the disease. Samples were seeded in Petri dishes containing PDA supplemented with antibiotics. Macroscopic and microscopic characteristics were recorded from colonies and structures produced in PDA, CLA and SNA media. The pathogenicity was evaluated by inoculating a suspension of conidia in the seed, and evaluating the percentage, index, and speed of germination, as well as the severity of the disease, survival analysis with Kaplan-Meier curves and the fit of epidemiological models of the progress of the disease was also determined. It was found that the fungal isolates that cause the damping-off problem in a pre-emergent and post-emergent way, obtained from seedlings of Pinus spp. in the forest nurseries of the state of Durango, were morphologically similar to isolates belonging to the F. fujikuroi and F. oxysporum species complexes. In addition, differences were observed in the severity of the disease produced by the isolates and they fitted the Gompertz model and the monomolecular model.
F. fujikuroi, F. oxysporum, Pinus spp., germination, survival.
Growing seedlings in a nursery is one of the most important activities of reforestation programs. To consider a seedling suitable for such programs, the National Forestry Commission (CONAFOR, for its acronym in Spanish) considers certain characteristics that must comply with the Mexican standard NMX-AA-170-SCFI-2016 (Secretaría de Economía, 2016). In Durango, in 2018, 8 291 544 plants were produced and 4 680.8 ha were reforested (CONAFOR, 2019a, 2019b), which corresponded to 9.6% of the area and 11% of the plant produced in the country, making it the state with the largest number of plants produced and the largest reforested area.
In a study conducted by Prieto et al. (2009), they reported that 100% of the nurseries studied presented problems caused by phytopathogenic fungi that cause damping-off.
The disease occurs in two ways: the first is called pre-emergent, where seeds in the process of germination are affected, and post-emergent, which attacks young seedlings that have not lignified the stem (Landis et al., 1989; Dar et al., 2011). This disease frequently occurred in seeds and seedlings of many forest species and other crops and is caused by a complex of microorganisms which includes oomycetes and fungi that see their development favored mainly by high moisture conditions (Fajardo-Mejía et al., 2016).
The most common genera of fungi that cause the damping-off problem are Fusarium and Rhizoctonia, of which Fusarium is the most important for its ubiquity (Dumroese and James, 2005). One of the main goals of phytopathology is to gain knowledge of the agents that cause diseases in cultivated plants and, through the application of appropriate techniques and tools, reduce the severity and incidence of diseases and thus reduce the losses caused by them (Solano-Bonilla and Brenes-Chacón, 2012).
The present study aimed to isolate fungi from pine seedlings with symptoms of the disease in forest nurseries in the state of Durango in order to identify them and characterize them by using brightfield optical microscopy and through the use of pathogenicity tests, and to determine the damage caused to the plant, the survival function, and the fit to epidemiological models.
Five nurseries distributed in the state of Durango were sampled in September 2018. INIFAP Nursery (23° 59’ 23.2” north latitude and 104° 37’ 26.2” west longitude, 1 879 masl), General Francisco Villa nursery (25° 58’ 19.7” north latitude and 104° 35’ 51.6” west longitude, 1 878 masl), nursery of the Faculty of Forest Sciences of the Juárez University of the State of Durango (24° 00’ 48.3” north latitude and 104° 41’ 03.2” west longitude, 1 899 masl) and two nurseries located in the municipality of Santiago Papasquiaro: General Emiliano Zapata Nursery (25° 03’ 40” north latitude and 105° 26’ 17” west longitude, 1 756 masl) and San Nicolás Nursery (24° 56’ 57” north latitude and 105° 25’ 02.8” west latitude, 1 806 masl).
The sampling consisted of extracting seedlings approximately 45 days old with damping-off symptoms; they were homogenized with phosphate buffer solution pH 7.2; serial decimal dilutions were performed and from dilutions 10-5 and 10-6, they were sown with the plate extension technique in potato dextrose agar (PDA) culture medium in duplicate. The plates were incubated at 28 °C for seven days in an incubator (Quincylab® 12-140). At the end of the incubation period, the aspect of fungal colonies was recorded (Gabrekiristos et al., 2020). To obtain axenic culture, consecutive transfers were performed by hypha tip in PDA.
To observe the overall microscopic characteristics, each isolate was cultured using the PDA microculture technique (Harris, 1986) and the way of inoculation proposed by Rosana et al. (2014). Microscopic observations were made with the 40x objective of the microscope (Leica® CME) and microconidia were measured with the AmScope® software version 3.7 (calibrated with a Euromex® AE 1110 micrometer with 0.01 mm reticle). The identification of each fungal isolate at the species complex level was carried out with the descriptions and keys reported by Leslie and Summerell (2006).
To observe the specific microscopy characteristics of the isolates, monosporic cultures were prepared with the adaptation of the technique applied by Obanor et al. (2013). Finally, mycelial discs from the edge of an isolated colony were transferred to Petri dishes with PDA, and they were incubated at room temperature for seven days with continuous light (Nelson et al., 1983) in SNA culture medium, and incubated at 25 °C for seven days (Leslie and Summerell, 2006), and also in carnation leaf agar (CLA) medium prepared with the technique described by Peluola et al. (2020) and incubated at 25 °C for seven days, with a photoperiod of 12 h of ultraviolet light (365 nm) and 12 h of white light (Britz et al., 2001).
The shape and size of microconidia, macroconidia, apical cell shape, base cell shape, length, width, dorsiventral curvature, and number of septa were recorded. One hundred measurements of length and width of macroconidia and microconidia produced by each isolate were taken, as suggested by Robles-Yerena et al. (2016); García-Díaz et al. (2017); Rentería-Martínez et al. (2019).
A nursery trial was carried out to verify the pathogenicity of the isolates (Koch’s postulates). The experiment was carried out in the forest nursery of INIFAP-Durango. We used rigid polyethylene containers with 98 cavities filled with a mixture composed of 50% peat moss, 25% expanded perlite, and 25% vermiculite with Osmocote® slow release (8 months) fertilizer of formulation 18-6-12 (N-P-K) at a dose of 4 g L-1 of prepared substrate.
The size of the experimental unit was 18 cavities; two treatments (absolute control and inoculated treatment) with five replications were evaluated, and the design applied was completely randomized. Each fungal isolate was evaluated on the pine species from which it was originally obtained. Prior to sowing, the seed received a flotation treatment, was disinfested in a commercial sodium hypochlorite solution, and rinsed with sterile distilled water and left to dry for 10 min (Aoudou et al., 2012).
The absolute control was sown after its pre-germination preparation by placing a seed 1 cm deep in each cavity and covered with substrate mixture. Regarding the seed for the inoculated treatment, the inoculation was carried out following the methodology of Morales-Rodríguez et al. (2018) with some modifications. Afterwards, one seed was deposited per cavity at a depth of 1 cm and covered with a mixture of substrate.
Daily germination counts were made from the first emerged seedling to 50 days after sowing (das). The variables evaluated were: germination percentage and germination index, with the following formulas (González-Zertuche and Orozco-Segovia, 1996).
And
Where: GP= germination percentage; sg= germinated seeds; ss= seeds sown; GI= germination index; ni= number of seeds germinated at day i, ti= number of days after sowing and N= total seeds sown; GS= germination speed; and t= germination time from sowing to germination of the last seed. Also, from the time the first seed germinated, seedlings killed by damping-off were counted daily for 180 days. These damaged seedlings were removed from the growth cavity for transfer to the laboratory and transfer to PDA culture medium.
From the developed colonies, they were transferred to plates prepared for microculture using the methodology of Harris (1986); Rosana et al. (2014) and were incubated at 25 °C for six days. Microscopic observations were made to corroborate the growth and verify Koch’s postulates and thus determine the percentage of damage in seedlings.
Germination and damage data were subjected to statistical analysis using the Wilcoxon test; the Log-Rank test was performed based on the Kaplan-Meier curves to determine the survival differences between treatments at 180 das. With the data on the percentage of damage of the isolates that presented significant differences compared to the control, disease progress curves and their fit to epidemiological, Gompertz, logistical, exponential, and monomolecular models were determined (Campbell and Madden 1990) in order to define the parameters of the equation and define the incidence of the disease. The statistical analysis was performed with the help of the InfoStat® software version 2017 (Di Rienzo et al., 2017) and graphs in SigmaPlot® version 10.0.
Seedlings of these species were found with the symptoms characteristic of damping-off: prostrate with generalized loss of turgor, constriction, and necrosis in the neck. Fungal isolates were obtained from these seedlings, and seven of the isolates showed macroscopically the colonial morphology of the genus Fusarium: dry surface and cotton-like mycelium and production of pale pink to purplish-pink pigments (Nelson et al., 1983; Watanabe, 2010).
The isolates obtained from the cultivated species during the sampling of the present study coincide with the findings of Menkis et al. (2006), in which fungi of the genus Fusarium were found colonizing roots of seedlings of Pinus sylvestris and Picea abies and no species of the genera Rhizoctonia, nor oomycetes of the genera Pythium and Phytophthora were found.
There were variations between these isolates, such as elevation of the colony (flat, umbonate, elevated, and convex) and the arrangement of the hyphae in the mycelium (inclined, slightly inclined, and erect). It was decided to separate these fungal isolates and assign them an ID according to these differences and the species of pine from which they were isolated. Isolate 1 was obtained from Pinus engelmannii (FPe1), isolate 2 from Pinus engelmannii (FPe2), isolate 3 from Pinus engelmannii (FPe3), isolate 4 from Pinus durangensis (FPd1), isolate 5 from Pinus arizonica (FPa1), isolate 6 from Pinus arizonica (FPa2), and isolate 7 from Pinus cooperi (FPc1).
When the isolates were cultured in SNA medium, the formation of ovoid-shaped microconidia without septa was observed, and these produced in false heads on monophialides and polyphialides (Figure 1). The average size of the microconidia was 6.54 μm long by 2.13 μm wide. Of these isolates, FPe1, FPe3 and FPd1 stand out for the development of coiled hyphae that did not develop in the other isolates. Isolates FPe2, FPa1, FPa2 and FPc1 produced terminal and intercalary chlamydospores when cultured in PDA, a detail that was not present in the other isolates.
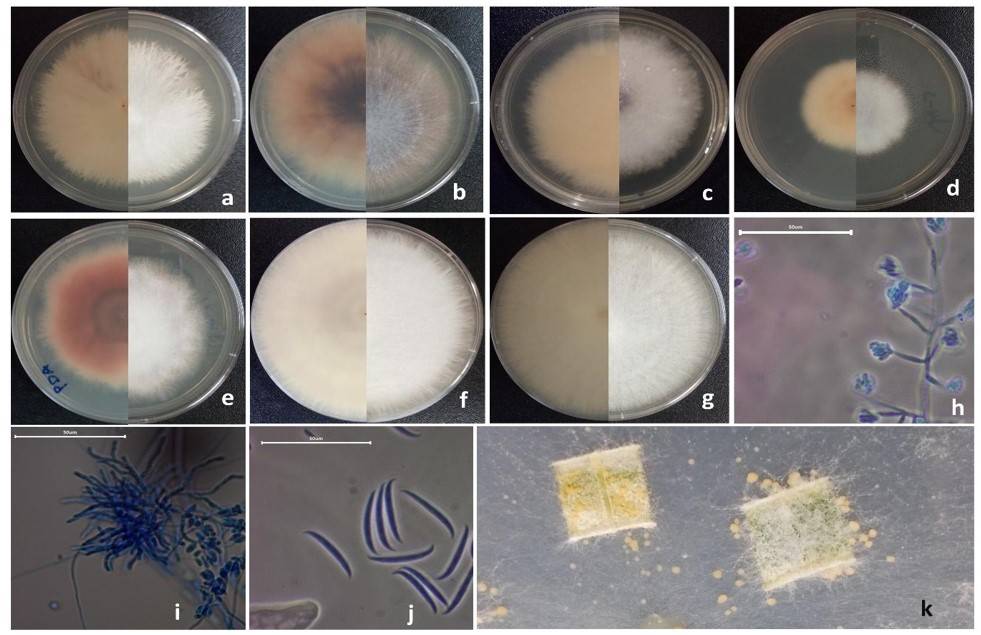
In the CLA medium, three isolates produced yellow to orange sporodochia (FPa1, FPa2, and FPc1), spindle-shaped macroconidia were visualized, with moderate dorsiventral curvature (Figure 1), with the base cell in the shape of a foot and the apical cell in a curved shape, with two and three septa in FPa1, FPa2, and three to four septa in FPc1. The size of the macroconidia of FPa1, FPa2 was 37.62 μm long by 3.3 μm wide, in FPc1 the size of the macroconidia was 23.63 μm long by 2.53 μm wide.
Given the macroscopic and microscopic characteristics of the isolates obtained and their morphological similarities with two Fusarium species complexes mentioned in the Fusarium laboratory manual (Leslie and Summerell, 2006), it was suggested that isolates FPe1, FPe3 and FPd1 corresponded to the characteristics of the Fusarium fujikuroi species complex and for isolates FPe2, FPa1, FPa2 and FPc1, they corresponded to the Fusarium oxysporum species complex.
The studies reported by Robles-Yerena et al. (2016); García-Díaz et al. (2018) are those that have been carried out in Mexico on the finding of Fusarium species that produce wilting problems in pine seedlings in the nursery stage and some are similar to some isolates in the present research.
After 1998, the description of new species increased, based on the recognition that several traditional morphospecies are species complexes composed of many species (Sumemerell and Leslie, 2011); for example, the species complex of F. fujikuroi contains 65 accepted species and includes some species that can only be identified based on their phylogenetic inference by molecular biology (Yilmaz et al., 2021); in addition, as mentioned by Crous et al. (2021) other lineages of Nectriaceae produce conidia with morphology similar to Fusarium and therefore the recognition of Fusarium species only with morphological traits is very complicated, which is why in the present work only concordances in the morphology of the obtained isolates to the F. fujikuroi (FFSC) and F. oxysporum (FOSC) complexes are suggested.
In most of the treatments inoculated with the isolates, there were no differences compared to the control in the variables of germination, germination index and germination speed, so it is confirmed that they did not cause pre-emergent damage and there was an average germination between 80 and 95% in them, only in the species P. arizonica, although there was no pre-emergence damage compared to the control, the average germination was 34 to 45%, possibly because the cultivation of this particular species is carried out in the north of the state of Durango, a location of more northern latitude than in the place where the nursery experiment was carried out.
In germination, there were only differences in a treatment inoculated in the species P. engelmannii (FPe3). In this, the germination percentage in the control was 90% and in the inoculated treatment it was 63%; that is, germination decreased by 27%, indicating that this isolate was the only one that caused pre-emergent damage and this was confirmed by the results obtained in the germination index analysis, since it also presented significant differences in this variable. Only in P. durangensis were there differences in the germination speed variable between the control and the inoculated treatment.
The accumulation of damaged seedlings was concentrated from 13 das to 40 das (week 2 to week 7). In the damage variable, the treatments inoculated with isolates that did not present differences compared to the control were FPe1, FPe2 and FPc1, so that they are considered to be the least virulent.
On the other hand, the treatments inoculated with the isolates that caused significant damage compared to the control, that is, the highest percentage of post-emergent damage in ascending order were: FPa1 (32.2%), FPd1 (60.7%), FPa2 (61.1%), and FPe 3 (100%). Isolate FPe3 (isolate 3) is considered to be the most virulent since it accumulated the largest number of damaged seedlings in the shortest time (Figure 2).
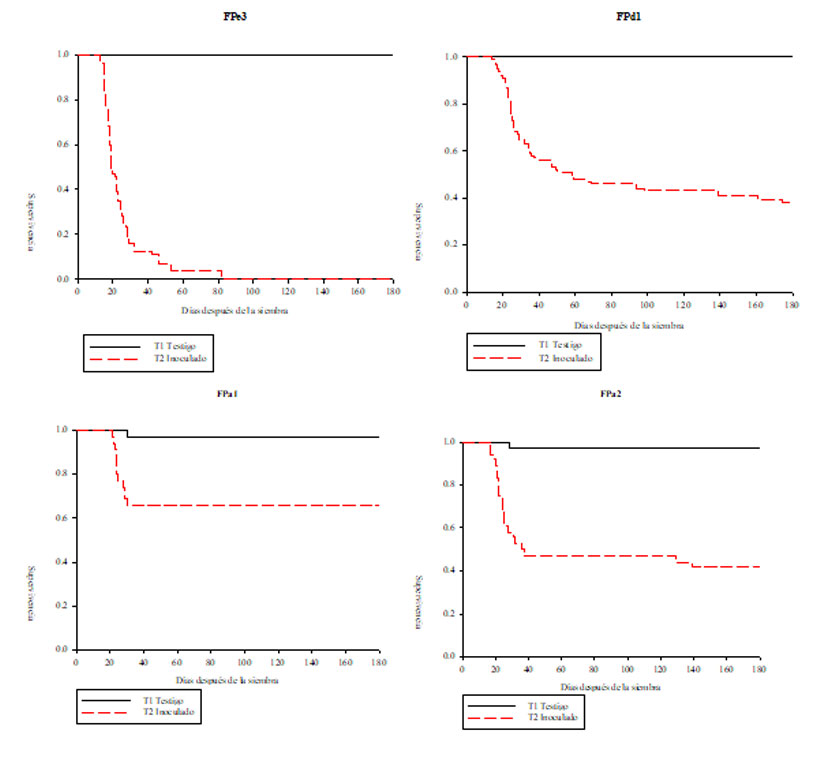
These results coincide with those described in other studies that have analyzed the pathogenesis of Fusarium in other pine species, such as those of Davydenko et al. (2018), who found that F. circinatum caused the death of almost all seedlings of Pinus sylvestris of different origins and F. oxysporum caused considerable damage, of 62-66% by day 21 after inoculation.
On the other hand, Mateos et al. (2017) found that F. oxysporum presented a higher index of damage compared to F. verticillioides in Pinus sylvestris and Pinus pinea. On the other hand, Lazreg et al. (2014) found that F. redolens and F. solani cause the most damage in pine seedlings, followed by F. tricinctum and F. oxysporum in Pinus halepensis.
In this essay, the first few weeks were key to pathogen development and seedling death. This period is the critical one to carry out the preventive and curative phytosanitary treatments necessary to stop the development of the disease. This is related to what was expressed by Davydenko et al. (2018), who indicated that greenhouses with plants infected with F. circinatum are the first point of entry of the disease and are a source of transmission of pine pitch canker in commercial forest plantations.
The epidemiological model that presented the best fit for the behavior of isolate FPe3 was the Gompertz model, represented by the expression damage= alpha*exp(-beta*exp(-gamma*DAS)), in which the parameters made a significant contribution (p< 0.05) and its parameters were alpha 98.36, beta 15.36 and gamma 0.15. Also for isolate FPd1, the best fit was shown by the Gompertz model and its parameters were alpha 55.78, beta 7.27 and gamma 0.08.
On the other hand, for the isolates obtained from P. arizonica, that is FPa1 and FPa2, the model that presented the best fit was the monomolecular model, represented with the expression damage=alpha*(1-beta*exp(-gamma*DAS)); for FPa1, the parameters were alpha 55.78, beta 7.27 and gamma 0.08; and for FPa2, the parameters were alpha 55.78, beta 7.27 and gamma 0.08.
In this context, Ocamb et al. (2002) mentioned that variation in Fusarium populations can be influenced during handling and sowing and this fungus can be introduced during seed storage or handling and they recommend pathogenicity tests to really know the potential for damage of Fusarium species isolated from seedlings.
It was possible to isolate fungi from seedlings with damping-off symptoms in nurseries in the state of Durango and these corresponded to isolates of the genus Fusarium from the F. fujikuroi and F. oxysporum species complexes according to their morphological traits. The most virulent isolates were obtained from Pinus engelmannii, P. durangensis and they fitted the Gompertz model, while the isolates obtained from P. arizonica fitted better the monomolecular model. It is necessary to carry out molecular identification of these isolates in order to know in detail the species of Fusarium that cause damping-off in pine seedlings that are produced in nurseries in the state of Durango.
This work was carried out thanks to the support of a postgraduate scholarship granted to the first author by CONACYT, also with the support provided by the projects PAICYT UANL CT 706-19, PAICYT, UANL CT 1187-20 of the Autonomous University of Nuevo León and INIFAP-Campo Experimental Valle del Guadiana. The authors would like to thank the reviewers who helped to improve the manuscript.
Britz, H.; Couhnho, T. A.; Gordon, T. R. and Wingfield, M. J. 2001. Characterizations of the pitch canker fungus, Fusarium circinatum, from Mexico. South African Journal of Botany. 67(4):609-614. https://doi.org/10.1016/S0254-6299(15)31190-X.
CONAFOR. 2019a. Comisión Nacional Forestal. Producción de plantas para reforestación. Sistema Nacional de Información Ambiental y de Recursos Naturales. México. http://dgeiawf.semarnat.gob.mx:8080/ibi-apps/WFServlet?IBIF-ex=D3-rforesta09-08&ibic-user=dgeia-mce&ibic-pass=dgeia-mce&nombreentidad=*&nombreanio=*.
CONAFOR. 2019b. Comisión Nacional Forestal. Superficie reforestada (hectáreas). Sistema Nacional de Información Ambiental y de Recursos Naturales. México . http://dgeiawf.semarnat.gob.mx:8080/ibi-apps/WFServlet?IBIF-ex=d3-rforesta09-06&ibic-user=dgeia-mce&ibic-pass=dgeia-mce&nombreentidad=*&nombreanio=*.
Crous, P. W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K. A.; Schroers, H. J.; Chaverri, P. and Thines, M. 2021. Fusarium: more than a node or a foot-shaped basal cell. Studies in Mycology. 98(0):1-184. https://doi:10.1016/j.simyco.2021.100116.
Dar, G. H.; Beig, M. A.; Ahanger, F. A.; Ganai, N. A. and Ahangar, M. A. 2011. Management of root rot caused by Rhizoctonia solani and Fusarium oxysporum in blue pine (Pinus wallichiana) through use of fungal antagonists. Asian journal of plant pathology. 5(2):62-67. https://doi.org/10.3923/ajppaj.2011.62.74.
Davydenko, K.; Nowakowska, J. A.; Kaluski, T.; Gawlak, M.; Sadowska, K.; García, J. M.; Diéz, J. J.; Okorski, A. and Oszako, T. 2018. A Comparative Study of the Pathogenicity of Fusarium circinatum and other Fusarium Species in Polish Provenances of P. sylvestris L. Forests. 9(9):1-13. https://doi.org/10.3390/f9090560.
Di Rienzo, J.; Casanoves, F.; Balzarini, M.; Gonzalez, L.; Tablada, M. y Robledo, C. 2017. InfoStat versión 2017. Córdoba, Argentina Universidad Nacional de Córdoba. http://www.infostat.com.ar.
Dumroese, R. K. and James, R. L. 2005. Root diseases in bareroot and container nurseries of the pacific northwest: epidemiology, management, and effects on outplanting performance. New Forests . 30(2-3):185-202. https://doi.org/10.1007/s11056-005-4422-7.
Fajardo-Mejía, M. A.; Morales-Osorio, J. G.; Antonio, G. and León-Peláez, J. D. 2016. Effect of plant extracts and growth substrates on controlling damping off in Pinus tecunumanii seedlings. Cerne. 22(3):317-324. https://doi.org/10.1590/01047760201622032150.
Gabrekiristos, E.; Teshome, D. and Ayana, G. 2020. Cultural, morphological and pathogenic variability among isolates of Fusarium oxysporum f. sp. capsici causing wilt of hot pepper in central rift valley, ethiopia. Journal of plant pathology & microbiology. 11(6):1-12. https://doi.org/10.35248/2157-7471.20.11.499.
García-Díaz, S. E.; Aldrete, A.; Alvarado-Rosales, D.; Cibrián-Tovar, D. y Méndez-Montiel, J. T. 2018. Trichoderma harzianum Rifai as a biocontrol of Fusarium circinatum Nirenberg & O’ Donnell in seedlings of Pinus greggii Engelm. ex Parl. in three substrates. Revista Chapingo Serie Ciencias Forestales y del Ambiente. 25(3):353-367. https://doi.org/10.5154/r.rchscfa.2018.12.088.
García-Díaz, S. E.; Aldrete, A.; Alvarado-Rosales, D.; Cibrián-Tovar, D.; Méndez-Montiel, J. T.; Valdovinos-Ponce, G. y Equíhua-Martínez, A. 2017. Efecto de Fusarium circinatum en la germinación y crecimiento de plántulas de Pinus greggii en tres sustratos. Agrociencia. 51(8):895-908. https://agrociencia-colpos.org/index.php/agrociencia/article/view/1334/1334.
Harris, J. L. 1986. Modified method for fungal slide culture. Journal of clinical microbiology. 24(3):460-461. https://doi.org/10.1128/jcm.24.3.460-461.1986.
Lazreg, F.; Belabid, L.; Sanchez, J. A. A.; Gallego, E. and Bayaa, B. 2014. Pathogenicity of Fusarium spp. associated with diseases of aleppo pine seedlings in algerian forest nurseries. Journal of Forest Science. 60(3):115-120. https://doi.org/10.17221/65/2013-JFS.
Mateos, E.; Olaizola, J.; Pajares, J. A.; Pando, V. y Diez, J. J. 2017. Influence of Suillus luteus on Fusarium damping off in pine seedlings. African journal of biotechnology. 16(6):268-273. https://doi.org/10.5897/AJB11.1164.
Menkis, A.; Vasiliauskas, R.; Taylor, A. S. F.; Stenström, E.; Stenlid, J. and Finlay, R. 2006. Fungi in decayed roots of conifer seedlings in forest nurseries, afforested clear cuts and abandoned farmland. Plant Pathology. 55(1):117-129. https://doi.org/10.1111/j.1365-3059.2005.01295.x.
Morales-Rodríguez, C.; Bastianelli, G.; Aleandri, M.; Chilosi, G. and Vannini, A. 2018. Application of Trichoderma spp. complex and biofumigation to control damping off of Pinus radiata D. Don caused by Fusarium circinatum nirenberg and O’Donnell. Forests . 9(7):1-16. https://doi.org/10.3390/f9070421.
Obanor, F.; Neate, S.; Simpfendorfer, S.; Sabburg, R.; Wilson, P. and Chakraborty, S. 2013. Fusarium graminearum and Fusarium pseudograminearum caused the 2010 head blight epidemics in Australia. Plant Pathology . 62(1):79-91. https://doi.org/10.1111/j.1365-3059.2012.02615.x.
Ocamb, C. M.; Juzwik, J. y Martin, F. B. 2002. Fusarium spp. and Pinus strobus seedlings: root disease pathogens and taxa associated with seed. New Forests . 24(1):67-79. https://doi.org/10.1023/A:1020589121134.
Peluola, C.; Hoesel, S. and Crutcher, F. 2020. Chlorine gas is an effective alternative to sterilize carnation leaves for Fusarium spp. identification. Journal of Microbiological Methods. 170(0):1-17. https://doi.org/10.1016/j.mimet.2020.105841.
Rentería-Martínez, M. E.; Guerra-Camacho, M. Á.; Ochoa-Meza, A.; Moreno-Salazar, S. F.; Meza-Moller, A. D. C. y Guzmán-Ortíz, J. M. 2019. Description and comparison among morphotypes of Fusarium brachygibbosum, F. falciforme and F. oxysporum pathogenic to watermelon in Sonora, México. Revista Mexicana de Fitopatología. 37(1):16-34. https://doi.org/10.18781/r.mex.fit.1808-1.
Robles-Yerena, L.; Leyva-Mir, S. G.; Cruz-Gómez, A.; Camacho-Tapia, M.; Nieto-Ángel, D. y Tovar-Pedraza, J. M. 2016. Fusarium oxysporum Schltdl. y Fusarium solani (Mart.) Sacc. causantes de la marchitez de plántulas de Pinus spp. en vivero. Revista Mexicana de Ciencias Forestales. 7(36):25-36. https://doi.org/10.29298/rmcf.v7i36.57.
Rosana, Y.; Matsuzawa, T.; Gonoi, T. and Karuniawati, A. 2014. Modified slide culture method for faster and easier identification of dermatophytes. Microbiology indonesia. 8(3):7-7. https://doi.org/10.5454/mi.8.3.7.
Solano-Bonilla, M. y Brenes-Chacón, D. 2012. Evaluación de métodos de curación de sustratos para la prevención del mal de talluelo. Revista Forestal Mesoamericana Kurú. 9(22):63-65. https://doi.org/10.18845/rfmk.v9i22.365.
Summerell, B. A. and Leslie, J. F. 2011. Fifty years of Fusarium: how could nine species have ever been enough? Fungal Diversity. 50(0):135-144. https:// 10.1007/s13225-011-0132-y.
Yilmaz, N.; Sandoval-Denis, M.; Lombard, L.; Visagie, C. M.; Wingfield, B. D. and Crous, P. W. 2021. Redefining species limits in the Fusarium fujikuroi species complex. persoonia molecular phylogeny and evolution of fungi. 46(1):129-162. https://doi.org/10.3767/persoonia.2021.46.05.