elocation-id: e3581
Floods are increasing worldwide because of the climate change. On the other hand, it is necessary to provide an adequate destination for the large amount of residue produced after banana harvesting. The objective of this study was to evaluate the effect of pseudostem residue on the recovery of banana seedlings subjected to waterlogging. The experiment was carried out in Registry, SP, Brazil, in a greenhouse, in the autumn and spring in 2021, using a design in divided plots with two factors and six repetitions. Waterlogging treatment was carried maintaining a layer of water in pots for three days (72 h). After drainage, pseudostem residue was added to half of the pots. seven days after the start of recovery, detected reduction of 20.3% in height and 28.7% in leaf area, accumulation of soluble carbohydrates in roots and leaves in autumn, reduction in chlorophyll, N, Mg and Zn contents, as well as the activity of POD, SOD and APX enzymes in roots and leaves. The residue proved to be beneficial after stress due to a 37.8% increase in the root area, recovery of Mg and Zn levels in root and leaf, prevention of elevation of activity of POD and SOD enzymes in leaves, APX in root. In the absence of stress, it reduced the activity of antioxidant enzymes; therefore, it should be used regardless of crop area be subject waterlogging.
i>Musa spp., banana waste, abiotic stress, flooding.
The frequency and intensity of floods are constantly increasing worldwide associated with global climate change being one of the most significant threats to food production and economic growth worldwide (Teoh et al., 2022). The reduction of soil oxygen levels caused by water excess, stops root growth and nutrient absorption in banana plants, causes death of juvenile roots, reduces photosynthesis and respiration rates and induces synthesis of ethylene as a response to the stress (Vargas et al., 2011).
Flooding also induces oxidative stress (Messchmidt et al., 2015), due to the formation of reactive oxygen species (ROS), resulting from the partial reduction of singlet oxygen (1O2), hydrogen peroxide (H2O2) and hydroxyl radical (•OH) (Seymen, 2021). If not eliminated, ROS can damage proteins, DNA and lipids and negatively affect several cellular functions (Messchmidt et al., 2015).
The antioxidant system formed by enzymatic and non-enzymatic components can control the overproduction of ROS under stress conditions and maintain the balance of homeostasis (Seymen, 2021).
The primary symptoms of oxygen deprivation in banana plant are wilting, yellowing, and reduced plant and fruit growth, even early yellowing during fruit transport (Robinson and Saúco, 2011). Timing of transplantation is also very important since young plants are more sensitive to flood than mature plants (Raman et al., 2022). Every 10 g plant-1 of dead functional roots due to flood results in a reduction of 40 to 140 banana boxes ha-1 year-1 (Serrano, 2003).
In a recent study, Teoh et al. (2022) proposed a model for the responses of banana plants to waterlogging based on anatomical, biochemical and gene expression study. According to them, under this stress, banana plants rapidly extended their adventitious roots, generate aerenchyma, and activate their antioxidant defense system in response to waterlogging.
On the other hand, pseudostem comprise about 60% of a banana plant's mass, represent a significant residue fraction in banana farms as they are cut down after each harvest (Castillo et al, 2023). Approximately, 220 t ha-1 year-1 of this residue are generated, constituting both an environmental problem related to soil toxicity and a high carbon footprint (Adsal et al., 2020). Pseudostem residue is rich in water, nutrients and polyphenols, among which phenols and flavonoids are powerful primary antioxidants or ROS scavengers (Sharma et al., 2017). It also contains hormones, amino acids, carbohydrates, and other metabolites that could contribute to increased growth.
Considering that the intensification of rainfall due to global warming will result in more flash flooding and seasonal flooding, the objective of this study was to evaluate the effect of pseudostem residue on the recovery of banana seedlings subjected to waterlogging under nursery conditions.
The experiments were carried out in the city of Registro, state of São Paulo, Brazil, in a greenhouse of Faculdade de Ciências Agrárias do Vale do Ribeira, UNESP, located at coordinates 24° 32’’ 08,57’’ south latitude and 47° 51’ 54,53’’ west longitude. The climate of the region according to the Köppen’s classification is Af, tropical rainy without dry season. Temperature data during the experiment were obtained from the Meteorological Station connected to the CIIAGRO System - IAC.
Seedlings of banana cv. Nanica (Musa spp. AAA) obtained by micropropagation were transplanted to pots of 10 dm3 containing 50% of soil, and 25% of commercial substrates Carolina and Vivatto. Fifteen days after transplanting, waterlogging treatment was applied maintained a 2 cm high layer of water for three days. Subsequently, water was drained of pots and added to half of them 0.5 kg of banana pseudostem residue, cut into pieces of 10 cm of height.
The adopted design was completely randomized in a factorial scheme and subdivided plots, with six replications. Located in the plot, the 2 x 2 factorial was constituted by the presence or absence of waterlogging and the presence or absence of residue. The autumn (May to July) and spring (October to December), seasons constituted the subplots, with a 36-day plant growth cycle in the two seasons.
The evaluations were performed seven days after the end of the waterlogging period. Initially, the height of the plant, the diameter of the pseudostem and the number of leaves were determined. The plants were carefully dug up and washed in water to remove soil adhering to the roots. After, separated into shoots and roots, and the material photographed. The images were submitted to the Image J software to estimate the leaf area and root area (Schneider et al., 2012). Small portions of fresh material were collected to determine the content of photosynthetic pigments and activity of antioxidant enzymes, the remainder was dried in an oven at 65 °C, followed by determination of the dry mass of the root and shoot. Portions of the dry material were ground determination of carbohydrate and nutrient contents.
Chlorophyll and carotenoid contents were determined in leaf discs after extraction with 80% acetone (v/v) (Wellburn, 1994). To determine the carbohydrate fractions in leaf (l) and root (r), 100 mg of crushed material were subjected to extraction with 80% ethanol at 75 °C for 20 min, repeating this procedure three times. The reducing sugars (RS) were quantified by the ninitrosalicylic acid method (Miller, 1959), while total soluble sugars (TSS) and starch, through reaction with anthrone (Hodge and Hofreiter, 1962). Non-reducing sugars (NRS) were quantified by the difference between TSS and RS. The nutrients contents (N, P, K, Ca, Mg, B, Fe, Cu, Mn e Zn) were determined according to Bataglia et al. (1983).
The enzymatic activity was determinate at the end of the stress period (day 0), at 7 and 14 days after the start of recovery. Fresh material (920 mg) of root and leaf was collected at 10:00 am and homogenized with 6 ml of 0.1 M potassium phosphate buffer, pH 6.8 and 10% PVPP (w/v). The macerate was filtered and centrifuged at 15000 rpm for 10 min at 4 °C. The extract obtained was used to determine the activity of superoxide dismutase (SOD) (Giannopolitis and Ries, 1977), peroxidase (POD) (Kar and Mishra, 1976), catalase (CAT) (Peixoto et al., 1999) and ascorbate peroxidase (APX) (Asada, 1984). To determine the specific activity of the enzymes, the Bradford (1976) was used to estimate soluble protein concentration.
The results were submitted to test of normality of Shapiro-Wilk. The analysis of variance was performing with the application of the F test (p<0.05) and the means of the treatments were compared using the Tukey test (p< 0.05). The software used for statistical analysis was SISVAR.
Waterlogging for three days induced varied responses in plant growth, but independent of the growing season (p< 0.01) (Figure 1). The negative impact was detected in the height and leaf area in the treatments waterlogging (W) and waterlogging with the addition of pseudostem residue (W+P), which even after seven days of the end of the stress showed lower mean values than control plants (C) and that received only pseudostem residue (P).
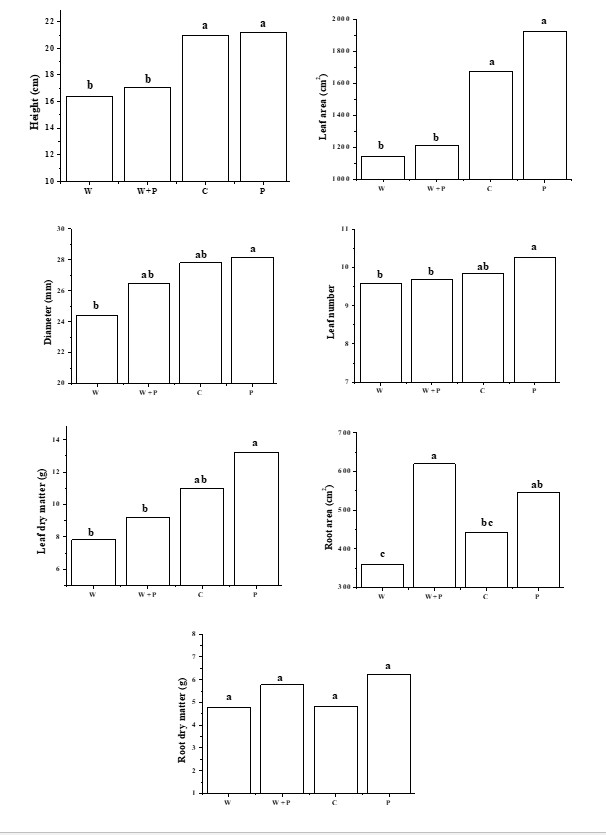
The diameter of the pseudostem, the number of leaves, dry mass of the leaves of the plants of the W and W+P treatments, despite being lower, did not differ from C. This is due the smaller effect of stress on these variables or yet the growth recovery after the drainage of the pots. The root area was reduced in W and higher than the control in W+P, indicating that the pseudostem residue was beneficial, despite the root dry mass not differing between treatments.
In general, in autumn, the leaf contents of total soluble sugars (TSS l), reducing sugars (RS l) and non-reducing sugars (NRS l) of the W and W+P treatments were higher than those of the C and P treatments, indicating that the stress caused an accumulation of soluble carbohydrates in the leaf tissues (Table 1). In spring, there was no difference between treatments for the different fractions of these carbohydrates. It is also noted that in autumn the mean values of the different fractions of soluble carbohydrates in leaf were higher than in spring, indicating an additional effect of the autumn season on carbohydrate metabolism.
In roots, the starch content was influenced only by the growing season, being higher in autumn (300.4) than in spring (162.44), regardless of treatments (Table 1). The root contents of total soluble sugars (TSS r) and reducing sugars (RS r) were influenced by the interaction of growing season and treatments, and in autumn these two fractions were higher in W and W+P compared to C, while the P treatment did not differ from control. In spring, TSS r and RS r were not influenced by treatments and exhibited lower mean values than in autumn. Non-reducing root sugars (NRS r), which includes sucrose, were not affected by season or waterlogging.
In autumn, the absolute minimum and máximum temperatures were 8.89 and 33.38 °C and the availability of total accumulated global radiation was only 369.10 MJ m-2, while in spring, the temperatures were 16.25 and 38.8 °C and the radiation accumulation was 544.13 MJ m-2. Robinson and Saúco (2011) point out that the temperature limits for dry matter gain in banana are between 8 and 15 °C, whose minimum limit was exceeded in half of the days of the crop cycle in autumn. There was also less radiation available in autumn compared to summer, which agrees with the fact that the average number of leaves, plant height and pseudostem diameter were lower in autumn than in spring (p< 0.05), regardless of treatments.
The accumulation of carbohydrates in both leaves and roots in autumn may be related to the lower rate of growth and development, which is especially greatly affected by temperature (Table 1). The highest levels of soluble carbohydrates in leaves and roots of plants subjected to waterlogging (W and W+P) in autumn are more related to the climatic condition than to the stress itself since no differences were detected between treatments in the spring.
Under waterlogging, soluble sugars produced in photosynthesis tend to accumulate in the leaves and are not translocated to the roots, where they are necessary to keep the glycolytic pathway functioning under conditions of oxygen limitation (Fante et al., 2010). In this study, higher average values of TSS r and RS r in the roots of plants from the W and W+P treatments in autumn reveal the lower use of carbohydrates as an energy source.
Regardless of the growing season, the total chlorophyll content in plants from treatments W (9.08 µg cm-2) and W+P (9.36 µg cm-2) were lower than plants from treatment P (10.27 µg cm-2) and control (C) (10.85 µg cm-2) (p< 0.05). Carotenoid contents were not influenced by the growing season or treatments, showing an average value of 0.09 µg cm-2. Although, Teoh et al. (2022) did not estimate the chlorophyll content and did not notice any visible damage to the plants, they observed that the leaves of the flooded banana plants showed yellowing after 24 days of waterlogging.
Although the pH and nutrient content of the soil used were adjusted for banana cultivation, there was a great variation in the foliar nutrient contents according to the treatments, at 7 days after the stress (Table 2). This also may reflect the reduction in the expansion of the leaf area in plants of the W and W+P treatments (Figure 1) or the recovery of plants after the waterlogging. For N, P and K, the effects of the treatments were independent of the growing season, while the other nutrients, except B, which was not affected, had responses dependent on the growing season and the treatments.
Leaf N contents, as well as Mg and Zn in spring, were negatively affected in waterlogging (W), when compared to control plants (C) (Table 2). However, plants from the W+P treatment, for had Mg and Zn contents that did not differ from the C, indicating that there was a recovery of the contents of these nutrients with the addition of residue, which did not happen for the N. The residue used contained 5, 0.5, 82, 8, 2.3 and 0.2 g kg-1 DM for N, P, K, Ca, Mg and S, and of 11, 3, 230, 168 and 8 mg kg-1 DM for B, Cu, Fe, Mn and Zn, respectively, and 45/1 of C/N ratio.
Plant’s N pool was reduced with waterlogging and that the use of residue was not enough to reduce the N deficit in the recovery period (Table 2). Due to the lower availability of energy in the form of ATP from aerobic respiration, the nutrient uptake by plants dramatically decreases, leading to plant starvation (Sasidharan et al., 2018). Despite the low N contents, the total protein content in leaves and roots at 7 days after stress did not vary between growing seasons and between treatments (p < 0.01), showing mean values, respectively, of 20.24 and 5.39 mg-1 g DM.
The leaf P content was affected only by the treatments, being lower in the control (C) and higher in plants of the W, W+P and P treatments, regardless of the growing season (Table 2). The higher levels of P in the leaves under stress (W and W+P) may be the result of the smaller expansion of the leaf area and in the W treatment, of the greater availability, absorption, and translocation due to the leachates. The higher concentration of K in the W treatment could also be the result of the lower leaf tissue growth, for this nutrient it was observed that the content in the W+P treatment did not differ from the control.
The foliar contents of Ca, Mg, Cu Fe, Mn, and Zn were influenced by the interaction between the growing season and treatments (Table 2). Ca, Fe and Cu showed similar response, higher contents in W and W+P treatments compared to C and P in autumn, reflecting the lower leaf tissue growth, while in spring there were no differences in treatments. Mg had an identical response to Ca and Fe in autumn, while in spring; the lowest mean values occurred in treatment W, compared to the other treatments that did not differ from each other. Higher Fe content in the leaves of plants subjected to waterlogging (W and W+P) in the autumn may also be the result of the greater availability and absorption of Fe in the reduced form (Fe+2) in an environment with O2 deprivation (Schmidt et al., 2013).
Mn did not vary between treatments in autumn but was higher in treatments that received residue in spring (W+P and P), indicating that its use contributes to its increment (Table 2). Zn also did not change in plants submitted to the different treatments in the autumn, and it was lower in plants of the F treatment in the spring in relation to the others. Leaf S content was not affected by growing season or treatment and by the interaction between these two factors.
In general, the most marked stress impact occurred in plants subjected to waterlogging (W) that exhibited lower foliar levels of N, as well as Mg, Mn and Zn in the spring, which were recovered with the addition of pseudostem residue (W+P), except for N (Table 2). The nutrient contents in the leaves were above the recommended for banana plants in the production phase, so it can be considered that the plants had a good nutritional status.
There was no effect of the growing season or treatments on the root P and K contents (Table 3). N and Zn content were influenced by the treatments independently of the growing season, unlike the contents of K, Ca, Mg, Fe, Mn, and B, which varied depending on the growing season, and treatments. N in the roots of treatment W (10.12 g kg-1) was lower than the control (14.52 g kg-1) and even with the addition of the residue (W+P) there was no complete recovery (11.98 g kg-1). The same response was found for Zn, with average values of 88.07, 96.23, 107.54 and 103.16 mg g-1, respectively for W, W+P, C and P.
Ca, Mg, Fe and Mn root contents tended to be higher in autumn, probably due to less use since climatic conditions were less favorable (Table 3). For Ca, in autumn, the highest content occurred in W, which was higher than the other treatments, while in spring, there were no differences between treatments. Mg content in autumn in roots of the W treatment did not differ from C and W+P, while in the spring, the W content was lower than in C, being recovered in W+P. The Fe content in the autumn in the roots of the treatment W, was superior to the control, and in the spring, W and C did not differ among themselves.
Mn content in the roots of treatment W was higher than that of C in autumn and spring (Table 3). Like Fe+2 and Mn+2 becomes more available with hypoxia in the soil because of waterlogging, which justifies the higher levels in the root under stress (Schmidt et al., 2013). B also exhibited higher root contents in plants under stress in autumn compared to C, whereas in spring, F and C would not differ. The levels of S and Cu in the roots were not influenced by any of the factors.
Interestingly, the nutrients affected by waterlogging (N, Mg and Zn) were the same in leaves and roots, suggesting a relationship. For N, the recovery of the contents with the addition of the pseudostem residue was incomplete in the leaves and roots, for Mg and Zn it was complete in roots and leaves.
In general, the enzymatic activity was not influenced by the interaction of waterlogging and residue. At the end of the stress, was detected reduction in the activity of peroxidase (POD) in the leaf and of superoxide dismutase (SOD) and ascorbate peroxidase (APX) in the root and leaf, in the two growing seasons in plants subjected to waterlogging (W) compared to C (Figure 2). However, there were no effects of growing season and treatments on catalase activity (CAT) in root and leaf, with average activities 0.68 and 1.38 µmoles H2O2 mg-1 protein, respectively.

SOD, POD, and CAT are the main detoxifying enzymes, which together with APX and glutathione reductase (GR) promote the elimination of reactive oxygen species (ROS) (Feng et al., 2015). In autumn, 7 days after the beginning of recovery (R7D), the activity of the same enzymes in W had already increased, reaching a level close to C, not differing from it. In the spring, however, at R7D, two distinct activity responses were observed in plants of the W treatment (Figure 2).
First, POD in the leaf and APX in the root with activity superior to the control, indicating an increase in the elimination of ROS, but that at 14 days after the onset of recovery (R14D) they exhibited normal activity. Second, at R7D in F, SOD activity in the root and leaf and APX in the leaf did not differ from the control, that is, an increase in activity after the end at a level close to the control and which was maintained at R14D.
It can be noted that for POD, SOD and APX in root in spring when enzymatic activities and the effect of stress were more intense, the presence of pseudostem residue after waterlogging (W+P) tended to prevent the increased activity compared to waterlogging (W) (Figure 2). At the same time, in spring, at R7D or R14D, the activity of the same enzymes in the leaves tended to be lower in the presence of the pseudostem residue and absence of stress (P).
The reduction in the activity of enzymes with waterlogging, regardless of the season, reflects the fact that aerobic respiration undergoes a rapid reduction with the deprivation of oxygen in the roots, as well as in energy production, as indicated by other studies with banana tree (Aguilar et al., 2003). Thus, initially due to the lower intensity of electron transport, the formation of ROS in the roots decreased in this period.
Stomatal conductance, photosynthesis, transpiration rates decreased during waterlogging periods in many species, including the reduction of photosynthetic pigments is a type of protective mechanism to maintain the photosynthetic structures of flooded plants to reduce sunlight absorption and prevent photo-oxidation (Du et al., 2012). In cherry tomatoes, after waterlogging (two days), the O2 - formation rate first decreased and then gradually increased (Liu et al., 2023).
After three days of waterlogging, only the APX activity was affected in the roots, while in the leaves, the activity of all enzymes was modified, depending on the time of cultivation (Figure 2). APX is involved in the formation of ascorbate, which acts as an antioxidant, and in the scavenging of H2O2 generated by SOD and POD (Feng et al., 2015).
In the root, the reduction of APX and maintenance of the activity of other enzymes under stress reflects the fact that aerobic respiration undergoes a rapid reduction with the deprivation of O2 in the roots, as well as in energy production, as indicated by other studies with banana plants (Aguilar et al., 2003). Therefore, due to the lower intensity of electron transport, the formation of ROS in the roots decreased in this period.
To FAPESP, for granting a scholarship (2019/18275-2) and to MM Mudas for donating seedlings
Teoh, E. Y.; Teo, C. H.; Baharum, N. A.; Pua, T. L. and Tan, B. C. 2022. Waterlogging stress induces antioxidant defense responses, aerenchyma formation and alters metabolisms of banana plants. Plants. 11(15):2052. https://doi.org/10.3390/plants11152052.