elocation-id: e3523
El género Fusarium afecta una amplia diversidad de cultivos hortícolas, generando importantes pérdidas económicas. Estos hongos son capaces de infectar a las plantas durante su desarrollo y aún después de cosecha. El uso de extractos vegetales bioactivos es una estrategia de control amigable al medio ambiente, no dañinas al consumidor, y de bajo costo. Los compuestos de origen vegetal han resultado ser promisorios en el control de enfermedades fúngicas. Cucurbita foetidissima es una cucurbitácea con alto contenido de metabolitos secundarios presentes en su raíz con poder fungicida. El presente trabajo se realizó durante 2017-2019 y se evaluó el efecto antifúngico de extractos radiculares de C. foetidissima frente a distintas especies de Fusarium bajo condiciones in vitro. Se obtuvieron extractos etanólicos y metanólicos radiculares de C. foetidissima. Se cuantificaron los metabolitos fenoles, flavonoides, terpenoides y saponinas. Se evaluó la capacidad antifúngica de los extractos sobre el crecimiento de Fusarium oxysporum, F. equiseti y F. solani in vitro. Los extractos metanólicos presentaron mayor concentración de fenoles, terpenoides y saponinas en comparación a los extractos etanólicos; estos últimos tuvieron mayor concentración de flavonoides. El extracto etanólico alcanzó mayor efecto antifúngico, inhibiendo entre 60 y 80% del crecimiento de las tres especies de Fusarium. El extracto etanólico radicular de C. foetidissima posee capacidad fungistática moderada para inhibir el crecimiento micelial in vitro de distintas especies de Fusarium, lo que sugiere el potencial de su uso para retrasar o atenuar la aparición de los síntomas asociados a fusariosis.
calabacilla silvestre, cucurbitaceas, fusariosis, metabolitos secundarios.
El género Fusarium incluye varias especies de hongos que causan enfermedades en diversos cultivos hortícolas, incluidos los principales cultivos alimentarios y comerciales como el trigo, la cebada, el maíz, tomate, plátanos, entre otros, generando importantes pérdidas económicas (DGSV-CNRF, 2020). Los posibles síntomas de enfermedades causadas por especies de Fusarium incluyen podredumbres de raíces o tallos, cancros, marchitez vascular, podredumbre de frutos o semillas y enfermedades de las hojas (Ekwomadu y Mwanza, 2023).
Para combatir dichas enfermedades, el control químico es la práctica más utilizada, ya que es relativamente económica, sencilla de aplicar y eficaz en comparación a otro tipo de control. Por años se han empleado de manera continua e indiscriminada los fungicidas sintéticos. Sin embargo, el uso excesivo de estos productos ha generado resistencia en los fitopatógenos (Kiiker et al., 2021), contaminación del medio ambiente y riesgos para la salud humana y animal (Tao et al., 2020).
Por esta razón se han buscado alternativas que sean amigables con el ambiente e inocuos al ser humano, como los compuestos vegetales que, además de presentar esas características, han demostrado tener potencial para el control de fitopatógenos. Los extractos vegetales pueden contener fenoles, antocianinas, terpenoides, flavonoides, taninos, lectinas, glucosinolatos, saponinas, entre otros, con actividad biológica (Mehmood et al., 2022).
Los resultados de Chacón et al. (2021) refieren que la presencia de compuestos fenólicos, aminas, flavonoides y terpenos en el aceite esencial de Piper auritum contribuyeron a la inhibición del crecimiento de F. oxysporum. Posteriomente, Isidro-Requejo et al. (2023) reportaron que el extracto de tomate, conteniendo fenoles y flavonoides, inhibió 100% el crecimiento micelial de F. oxysporum, F. graminearum y F. verticillioides.
Los extractos de hojas de Stevia rebaudiana, que contienen la saponina austroinulina, inhibieron el crecimiento de F. oxysporum en plantas de tomate (Ramírez et al., 2020). En un estudio similar, las saponinas aisladas de la quinoa (Chenopodium quinoa) redujeron significativamente la incidencia del marchitamiento por F. oxysporum en semillas y plántulas de tomate (Zhou et al., 2023).
Cucurbita foetidissima, conocida comúnmente como calabacilla loca, es un vegetal xerófito y silvestre de las regiones semiáridas de México y del sur de EE. UU. Por años se usó como alimento, medicamento y desinfectante (Gómez et al., 2019). Es hospedera asintomática de F. oxysporum (Apodaca-Sánchez et al., 2004), se le atribuyen propiedades medicinales (Mukherjee et al., 2022), y antifúngicas (Mejía-Morales et al., 2021).
Además, presenta varias cucurbitacinas y glicósidos de cucurbitacinas en el fruto, grupo de compuestos triterpénicos que proporcionan el sabor amargo y poseen actividad antimicrobiana, antifúngica y citotóxica (Mejía-Morales et al., 2021). Rangel-Guerrero et al., 2018) reportan que extractos obtenidos de fruta de C. foetidissima inhibieron entre el 94.3 y 100% el crecimiento de colonias de Rhizoctonia sp., mientras que en Fusarium sp. y Phytophthora sp., presentaron bajos porcentajes de inhibición (25.7-46.7%; 25.3-42.2%, respectivamente). Este efecto antifúngico puede deberse a la presencia de triterpenos tetracíclicos observados en esta planta.
Existen escasos estudios sobre metabolitos secundarios presentes en la raíz de C. foetidissima y su posible actividad antifúngica. Por lo anterior, el objetivo de este trabajo fue determinar e identificar los metabolitos secundarios presentes en extractos metanólicos y etanólicos de raíz de C. foestidissima y su actividad antifúngica in vitro sobre distintas especies de Fusarium. Lo anterior, con el fin de establecer las bases que posibiliten el desarrollo de un fungicida natural inocuo y eficaz para el control de la fusariosis.
En el 2017 se colectaron en forma aleatoria raíces de plantas adultas de C. foetidissima en etapa fenológica de fructificación (verano), procedentes de lomerío con suelos franco-arcillosos ligeramente alcalinos y salinos del municipio de Ciudad Cuauhtémoc, Chihuahua, con coordenadas 28° 44’ 763’’ latitud norte, 106° 82’ 887’’ longitud oeste y a 2 060 msnm. Las muestras se transportaron vía terrestre a las instalaciones del Centro de Investigación en Alimentación y Desarrollo, AC (CIAD), ubicado en Hermosillo, Sonora y almacenadas en refrigeración hasta su desinfección.
Las raíces se enjuagaron con abundante agua corriente para retirarles el excedente de suelo y posteriormente fueron desinfectadas con NaClO al 6%. Se cortaron en trozos pequeños, se secaron empleando una estufa de convección de aire forzado (Yamato DX-600, Japón), a 60 °C durante 72 h y la materia seca obtenida se pulverizó en un molino eléctrico (HB 8365, China).
El material deshidratado se maceró con dos solventes distintos (metanol y etanol, Sigma-Aldrich, USA) a una proporción de 1:3 p/v (Chapagain et al., 2007). Posteriormente se mantuvieron en agitación continua a 120 rpm en un agitador orbital (Environ Shaker, USA) durante ocho días a temperatura ambiente. El extracto se obtuvo por filtración al vacío y se concentró en un rotoevaporador (Yamato RE500, Japón) hasta sequedad (Chapagain et al., 2007). El pellet resultante se resuspendió en 25 ml de metanol (MEA) o etanol (EEA) y después se almacenó a -80 °C en tubos revestidos con aluminio para su posterior análisis. El proceso de extracción se realizó por triplicado.
Con el fin de establecer los grupos predominantes de metabolitos secundarios en los extractos metanólicos y etanólicos de la raíz de C. foetidissima, se aplicaron las pruebas presuntivas reportadas por García-Granados et al. (2019). Se utilizó la prueba del FeCl3 para detectar la presencia de fenoles (García-Granados et al., 2019). Se agregaron dos a tres gotas de FeCl3 al 12.5% a una alícuota de cada extracto. El cambio de color verde oscuro a azul índigo indicó la presencia de fenoles. En flavonoides se utilizó la prueba de Shinoda (García-Granados et al. (2019). Se tomó 1 ml de cada extracto, se agregó 0.1 g de magnesio y se calentó a 60 °C. Posteriormente se añadieron unas cuantas gotas de HCl. El cambio de coloración a naranja, rojo, rosa, azul o violeta indicó prueba positiva.
Los fitoesteroles y triterpenos se detectaron con la prueba de Libermann-Burchardt (García-Granados et al., 2019). Se tomó una muestra de 500 mg de cada extracto y se disolvió en 10 mL de cloroformo. Se tomaron 5 ml y se adicionaron 0.25 ml de anhídrido acético más dos gotas de H2SO4 concentrado. La formación de color verde indicó prueba positiva para fitoesteroles y un color rosa a morado indicó prueba positiva para triterpenos. Además, se determinó saponinas mediante la prueba de espuma. Se tomó una muestra de 500 mg y se añadieron 10 ml de agua destilada. Se agitó durante 30 s en vórtex y se dejó reposar por 15 min. La prueba fue positiva si la altura de la espuma fue de 5 mm o mayor.
Con base en los resultados obtenidos en el apartado anterior, se procedió a cuantificar el contenido de fenoles totales, flavonoides, terpenoides y saponinas. Se preparó muestra liofilizada utilizando trozos pequeños de la raíz limpia y desinfectada como se describió anteriormente, se congelaron a -80 °C por 24 h y se liofilizaron en un liofilizador (Yamato DC801, USA). Se preparó un extracto utilizando 0.25 g de muestra liofilizada, se añadió 5 ml etanol, se homogenizó en frío (Ultraturax IKA, Alemania) por 2 min a 3 000 rpm (Treviño et al., 2012).
Se decantó y colectó el sobrenadante, se añadió 5 ml de solución extractora (etanol o metanol) y se repitió la operación, se sonicó 60 min a 25 °C. Posteriormente se dejó reposar 2 h, se tomaron 1.5 ml y se centrifugó por 15 min a 13 000 rpm, el sobrenadante se separó y se utilizó en los siguientes análisis.
El contenido de fenoles totales se determinó con la técnica de Singleton et al. (1999) con ligeras modificaciones. Se preparó una mezcla de reacción con 40 µl del extracto, 60 µl de etanol, 0.4 ml de agua, 5 ml de NaOH, y 0.5 ml de la solución de Folin-ciocalteu (Sigma-Aldrich, USA). Después de 0.5 a 1 h de reposo, se midió la absorbancia a 765 nm. Se cuantificó con base a una curva de calibración, utilizando como estándar el ácido gálico (AG). Los resultados se reportaron como miligramos de ácido gálico equivalentes por gramo de extracto (meq AG g-1).
Los flavonoides se determinaron con la técnica de Zhishen et al. (1999) con ligeras modificaciones. La reacción de los extractos con el AlCl3 produce un color rosa intenso que se midió a una longitud de onda de 415 nm. Se determinó su contenido con base al estándar quercetina. Los resultados se expresaron como miligramos de quercetina equivalentes por gramo de extracto (meq Q.g-1).
Los terpenoides se determinaron con la técnica de Ghorai (2012) con ligeras modificaciones. Se tomó una alícuota del extracto y se le añadió 100 µl de H2SO4 concentrado formando un complejo de color rojo-marrón. Se midió la absorbancia a una longitud de onda de 538 nm, y cuantificó con base al estándar linalol. Los resultados se expresaron en mg de linalol g-1 de extracto.
Las saponinas totales se determinaron con la técnica de Helaly et al. (2001). Al extracto se le añadió 5 ml de H2SO4 al 72% y se midió la absorbancia a 435 nm. Se cuantificó de acuerdo el estándar ácido oleanólico. Los resultados se reportaron como mg equivalentes de ácido oleanólico g-1 de extracto. Todos los reactivos y estándares utilizados en las distintas técnicas fueron marca Sigma-Aldrich. Todas las determinaciones descritas en esta sección fueron cuantificadas por triplicado empleando un espectrofotómetro UV-Vis (Hach DR 5000, Alemania).
Se utilizaron tres especies de Fusarium (F. oxysporum, F. solani y F. equiseti) previamente aisladas de frutos de melón cantaloupe. Los hongos se sembraron en cajas petri con agar papa dextrosa (PDA, Difco, USA) durante 10 días a 26 °C, para determinar las características de colonia: textura, color y conidiación. Se observaron sus características microscópicas utilizando un microscopio óptico (Binocular Primo Star Carl Zeiss). Los hongos se identificaron a especie de acuerdo con las claves taxonómicas reportadas por Leslie y Summerell (2008) y Tousson y Nelson (1976). Las colonias se conservaron en refrigeración (4 °C).
La capacidad de los extractos de C. foetidissima para inhibir el crecimiento micelial de las especies de Fusarium se evaluó in vitro por medio del método de pocillo (Pérez-Delgado y Vallejos-Campos, 2019). Se evaluaron los siguientes tratamientos: extracto etanólico de raíz de C. foetidissima (EEA), extracto metanólico de raíz de C. foetidissima (MEA), testigo etanol como solvente de extracción (TE) y testigo metanol como solvente de extracción (TM). En placas petri de 90 mm con PDA se perforaron cinco pozos de 6 mm de diámetro ubicados a 23 mm del centro de la placa. En cada pocillo se colocaron 100 µl del extracto según su tratamiento. En cada placa petri se colocaron discos de micelio (6 mm de diámetro) de cada especie de Fusarium. Se incluyó un control negativo con 100 µl de H2O destilada estéril (testigo).
Se realizaron tres repeticiones por cada uno de los tratamientos. La capacidad inhibitoria de los extractos se determinó con base en el crecimiento micelial del hongo, el cual fue calculado midiendo con un vernier el diámetro del crecimiento micelial cada 24 h durante 10 días. La capacidad inhibitoria se expresó en porcentaje de inhibición empleando la ecuación propuesta por Chapagain et al. (2007).
Donde: AC= promedio del área control; at = área individual del tratamiento. Con el área obtenida se calcularon la velocidad de crecimiento (VCr= área tiempo, mm2 día-1) y eficiencia de crecimiento (ECr = mm día-1).
El experimento se repitió dos veces y los datos se analizaron mediante un diseño completamente al azar. Para realizar el análisis estadístico del porcentaje de inhibición, los datos originales se transformaron por la fórmula arcsen√%. Se realizó un análisis de varianza (Anova) y la prueba de Tukey-Kramer para determinar diferencias entre medias a una probabilidad del 0.05%. Se utilizó el programa estadístico NCSS versión 2017.
Las pruebas presuntivas revelaron la presencia de fenoles, flavonoides, terpenoides y saponinas, los cuales han sido descritos como antifúngicos en diversos tejidos vegetales (Arif et al., 2009). Los resultados confirman lo reportado en hojas y frutos de C. foetidissima por Macías et al. (2009), quienes encontraron la presencia de fenoles, terpenoides y saponinas. Además de lo obtenido en hojas y frutos también en raíces encontraron abundancia de estos compuestos junto con compuestos triterpénicos (cucurbitacinas) y saponinas (Mejía-Morales et al., 2021).
En el Cuadro 1, se muestran las concentraciones de los compuestos cuantificados en los extractos. El solvente extractor donde se observó una mayor concentración de saponinas fue metanol, lo que concuerda con lo reportado en extractos de semillas de Camelia sativa L. donde se obtuvo hasta un 50% más concentrado de fenoles totales en extractos metanólicos que en etanólicos (Gupta y Kumar, 2017), mientras que los extractos etanólicos únicamente en flavonoides se obtuvo mayor concentración. Las saponinas fueron los compuestos más abundantes en los extractos metanólicos. Diversos autores describen a la planta de C. foetidissima como una planta altamente productora de saponinas y otros compuestos triterpénicos como las cucurbitacinas (Kaushik et al., 2015; Mejia-Morales et al., 2021).
Los extractos radiculares de C. foetidissima presentaron actividad antifúngica sobre las tres especies de Fusarium evaluadas. En la Figura 1, se muestra el efecto de los extractos en la inhibición del crecimiento micelial de Fusarium sp. bajo condiciones in vitro. Al comparar los tratamientos mediante la prueba de Tukey (0.05) se observó que los extractos metanólicos y etanólicos redujeron significativamente (p≤ 0.05) el crecimiento micelial de F. oxysporum (Figura 1A) desde el primer día de evaluación.
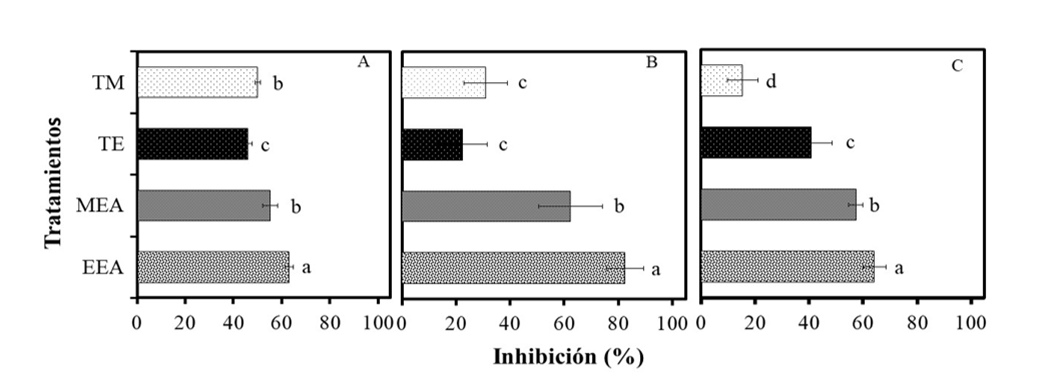
La aplicación del extracto etanólico (EEA) redujo significativamente el crecimiento del hongo, logrando el máximo porcentaje de inhibición el día 5 con 63%; mientras que el porcentaje de inhibición fue ligeramente menor en el extracto metanólico (MEA). El tratamiento EEA inhibió 15% más que los tratamientos testigo (TM y TE) y 9% más en comparación al extracto metanólico (MEA).
Ambos extractos tuvieron mayor capacidad inhibitoria en F. equiseti (Figura 1B), en comparación a las otras dos especies de Fusarium. La aplicación del EEA inhibió en mayor proporción el crecimiento micelial superando el 80% de inhibición, mientras que el extracto MEA solo inhibió el 60% el crecimiento de este hongo.
El porcentaje de inhibición en la especie de F. solani (Figura 1C) mostró un comportamiento similar a F. oxysporum. De nuevo, el tratamiento EEA alcanzó el 65% de inhibición, 10% más que el extracto MEA. La actividad inhibitoria de los extractos de raíz de C. foetidissima observados en el presente estudio es superior a la reportada por Rangel-Guerrero et al. (2018) quienes observaron que extractos metanólicos de fruto de C. foetidissima provenientes de distintas localidades de México, inhibieron el crecimiento de Fusarium sp., entre 25.7 y 46.7% a los 5 días de incubación a 27 °C.
Esto sugiere que la raíz de C. foetidissima contiene metabolitos secundarios con mayor actividad antifúngica, que el fruto de esta cucurbitácea. Es importante destacar que el extracto etanólico tuvo mayor capacidad inhibitoria con respecto al extracto metanólico, lo cual puede deberse al alto contenido de flavonoides que se registró en este estudio.
Hallazgos como los de Ammar et al. (2013), han identificado a los flavonoides presentes en plantas de Tephrosia apollinea L. como responsables de la actividad antimicrobiana sobre otros microorganismos fitopatógenos. Al Aboody y Mickymaray (2020) señalan que los flavonoides inhiben el crecimiento fúngico a través de varios mecanismos de acción, provocando la alteración de la membrana plasmática, la inducción de la disfunción mitocondrial, la inhibición de la formación de la pared celular, inhibición de la división celular e inhibición de la síntesis de ARN y proteínas.
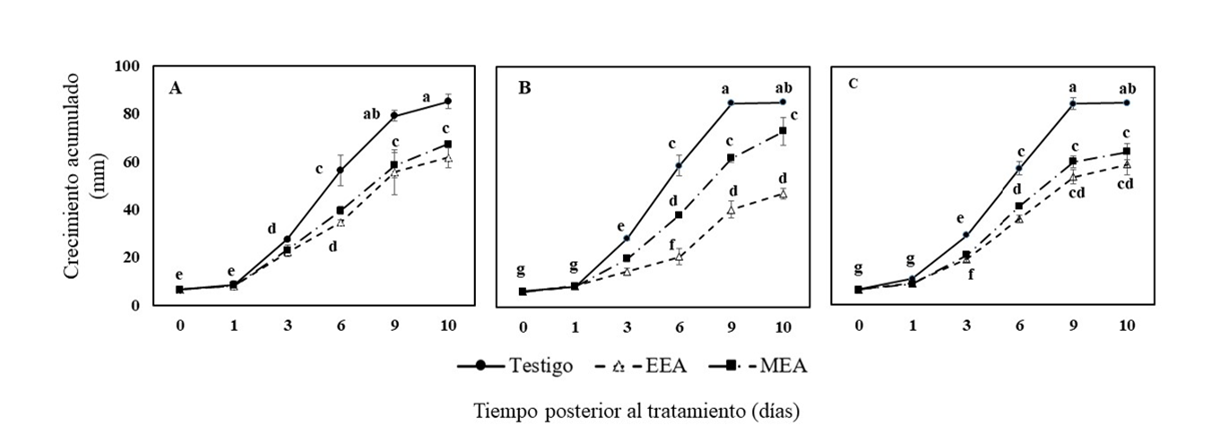
Con respecto a la cinética de crecimiento, las tres especies de Fusarium expuestas a extractos de C. foetidissima presentaron un patrón de crecimiento similar. En la Figura 2 se puede apreciar que tanto F. oxysporum (Figura 2A) como F. solani (Figura 2C) no presentaron diferencias significativas en su crecimiento al ser expuestas a ambos extractos, pero si fue menor significativamente (p≤ 0.05) con respecto al testigo.
En cambio, F. equiseti (Figura 2B) al ser expuesto al extracto etanólico, presentó un crecimiento menor con respecto a los otros tratamientos y testigo. Esto sugiere que F. equiseti es más sensible a los compuestos bioactivos extraídos con etanol con respecto a las otras especies de Fusarium evaluadas en el presente estudio.
Por otro lado, los resultados obtenidos sobre la eficiencia de crecimiento de las especies de Fusarium en respuesta a los extractos de C. foetidissima, se presentan en la Figura 3. Se pudo observar que la mayor eficiencia se encontró en el segundo día, con valores entre 1.6 y 2.09 mm día-1 para F. oxysporum (Figura 3A) y F. solani (Figura 3B). A partir de este día la eficiencia disminuyó constantemente hasta alcanzar el valor mínimo de su crecimiento (0.21 mm día-1). En el caso de F. equiseti (Figura 3C) la eficiencia de crecimiento fue menor con respecto a las otras especies de Fusarium.
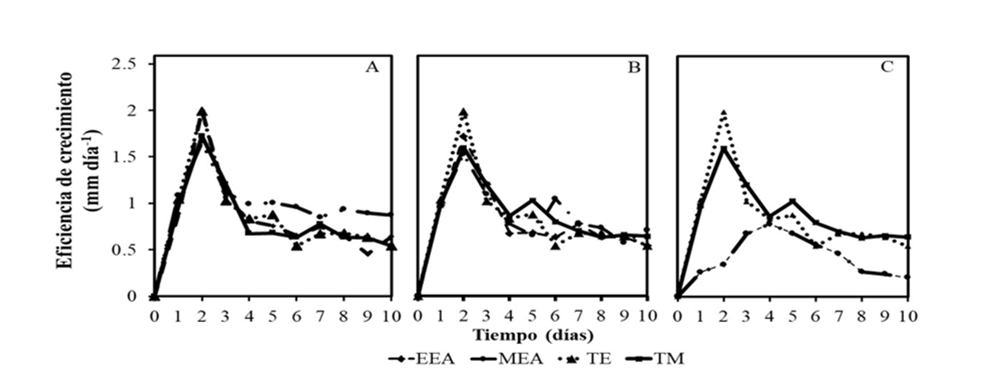
El comportamiento observado sobre el incremento en la eficiencia de crecimiento hasta un pico máximo seguido de una disminución de ésta podría deberse a la fotolabilidad de los compuestos fenólicos presentes en los extractos (Mohamed et al., 2020). Por otro lado, la capacidad del patógeno a producir y activar enzimas detoxificantes que inhiben la acción antifúngica de los compuestos (Mahgoub et al., 2023), favorece su crecimiento en condiciones adversas.
Se detectaron los metabolitos secundarios fenoles, flavonoides, terpenoides y saponinas en los extractos etanólicos y metanólicos de las raíces de C. foetidissima. El extracto metanólico presentó mayor concentración de fenoles, terpenoides y saponinas, mientras que el extracto etanólico presentó mayor concentración de flavonoides. El extracto etanólico de raíz de C. foetidissima presentó mayor capacidad antifúngica para inhibir el crecimiento micelial in vitro de F. oxysporum, F. equiseti y F. solani, lo que representó una alternativa promisoria que permitió retrasar o atenuar la aparición de los síntomas asociados a fusariosis.
Chacón, C.; Miranda-Granados, J.; Ruiz-Lau, N.; Lagunas-Rivera, S.; Ruíz-Valdiviezo, V. M. and Gutiérrez-Miceli, F. A. 2021. In vitro antifungal activity and chemical composition of Piper auritum Kunth essential oil against Fusarium oxysporum and Fusarium equiseti. Agronomy. 11(6):1-13. Doi:10.3390/ agronomy11061098.
DGSV-CNRF. 2020. Podredumbre de raíces por Fusarium spp. (Hypocreales: Nectriaceae) en maíz. SADER-SENASICA. Dirección General de Sanidad Vegetal-Centro Nacional de Referencia Fitosanitaria. Ficha técnica. 15 p. Tecámac, Estado de México. https://www.gob.mx/cms/uploads/attachment/file/600884/Podredumbre-de-ra-ces.pdf
García-Granados, R. U.; Cruz-Sosa, F.; Alarcón-Aguilar, F. J.; Nieto-Trujillo, A. y Gallegos-Martínez, M. E. 2019. Análisis fitoquímico cualitativo de los extractos acuosos de Thalassia testudinum Banks ex Köning et Sims de la localidad de Champotón, Campeche, México, durante el ciclo anual 2016-2017. Polibotánica. 1(48):151-168. Doi: 10.18387/polibotanica.48.12.
Isidro-Requejo, L. M.; Márquez-Ríos, E.; Toro-Sánchez, C. L.; Ruiz-Cruz, S.; Valero-Garrido, D. and Suárez-Jiménez, G. M. 2023. Tomato plant extract (Lycopersicon esculentum) obtained from agroindustrial byproducts and its antifungal activity against Fusarium spp. Frontiers in Sustainable Food Systems. 7(11):1-8. Doi: 10.3389/fsufs.2023.1323489.
Mahgoub, S. A.; Qattan, S. Y. A.; Salem, S. S.; Abdelbasit, H. M.; Raafat, M.; Ashkan, M. F.; Al-Quwaie, D. A.; Motwali, E. A.; Alqahtani, F. S. and Abd El-Fattah, H. I. 2023. Characterization and biodegradation of phenol by Pseudomonas aeruginosa and Klebsiella variicola strains isolated from sewage sludge and their effect on soybean seeds germination. Molecules. 26-28(3):1-19. Doi: 10.3390/molecules28031203.
Mejía-Morales, C.; Rodríguez-Macias, R.; Salcedo-Pérez, E.; Zamora-Natera, J. F.; Rodríguez-Zaragoza, F. A.; Molina-Torres, J.; Délano-Frier, J. P. and Zañudo-Hernández, J. 2021. Contrasting metabolic fingerprints and seed protein profiles of Cucurbita foetidissima and C. radicans fruits from federal plants sampled in Central Mexico. Plants. 10(11):1-22. Doi: 10.3390/plants10112451.
Rangel-Guerrero, J. M.; Flores-Benitez, S.; Cadena-Iñiguez, J.; Morales-Flores, F. J. and Trejo-Téllez, B. I. 2018. Extracts of Cucurbita foetidissima (Kunth) fruits inhibit the growth of phytopathogens of agricultural interest. Agroproductividad. 11(10):107-115. Doi.org/ 10.32854/agrop.v11i10.1253.