La estabilidad de la población de tallos permite analizar las variaciones en las tasas de aparición, muerte y supervivencia, por lo que, el objetivo de esta investigación fue determinar el efecto de la intensidad de cosecha (15 y 25 cm) en la estabilidad de la población de tallos de Urochloa híbrido Mulato II, Cayman y Mavuno. El estudio se realizó de marzo de 2020 a febrero de 2021 en Güémez, Tamaulipas, México. En praderas establecidas se delimitaron 18 parcelas experimentales de 3 x 3 m. En cada unidad experimental se colocaron dos áreas de 100 cm2 para seleccionar la población inicial de tallos y cada cuatro semanas contar los tallos muertos e hijuelos presentes. Se encontraron diferencias entre las intensidades y fechas de muestreo para las variables densidad de tallos, tasa de ahijamiento, e índice de estabilidad (p< 0.05). La intensidad de corte de 15 cm generó la mayor densidad de tallos con 2 545, 2 996 y 2 200 tallos m2 en Mulato II, Cayman y Mavuno, respectivamente, como resultado de tasas de ahijamiento mayores a 12% sin presentar diferencias en las tasas de muerte y porcentaje de sobrevivencia. La intensidad de corte de 15 cm promedió índices de estabilidad de 0.98 y 0.96 en Mulato II y Mavuno, mientras que, en Cayman se presentó estabilidad negativa (0.93). La intensidad de corte de 15 cm permitió mantener la estabilidad de la población de tallos de los híbridos Urochloa Mulato II y Mavuno a través del tiempo.
intensidad, persistencia, supervivencia
En México la ganadería extensiva se realiza en pastizales de regiones semiáridas (Velázquez et al., 2015), donde, la principal fuente de alimento son los matorrales, pastos y malezas (Romero-Paredes y Ramírez, 2003), con especies vegetales nativas e introducidas (Elizondo y Boschini, 2002; Quero et al., 2007); sin embargo, la distribución de la precipitación pluvial en estos sitios es errática, la cual, oscila entre 350 y 600 mm anuales para la zona semiárida y menor a 350 mm para la zona árida (Tarango, 2005), aunado a ello, los periodos prolongados de sequía producen un decremento en la producción animal (Cuartas et al., 2014), debido a que las plantas detienen su desarrollo y por ende, disminuye la disponibilidad de forraje (Carmona-Muñoz et al., 2003).
En este contexto, en el estado de Tamaulipas se han introducido especies del género Urochloa, con rendimientos de hasta 10.13 t ha-1 de materia seca acumulada en la época de máxima precipitación que corresponde a los meses de mayo a octubre en contraste a 1.36 t ha-1 presentes en los meses de noviembre a abril con un programa de manejo a intervalos de ocho semanas entre cortes, rendimientos superiores a Pennisetum ciliare H-17 (Garay-Martínez et al., 2018).
Los híbridos derivados del género Urochloa han sido desarrollados con características que los hace tolerantes al déficit de humedad en el suelo, incluso hasta llegar a marchitez permanente sin presentar cambios en la turgencia foliar y obtener mayor biomasa foliar en comparación a sus progenitores U. decumbens y U. brizantha (Carmona-Muñoz et al., 2003). No obstante, la respuesta de las especies en las características morfológicas y estructurales depende de diversos factores como edad de la planta, su composición genética, las condiciones climáticas y el manejo agronómico (Garay-Martínez et al., 2018).
Con el manejo agronómico, además de buscar rendimiento y valor nutritivo, se busca incrementar el tiempo de utilización del área establecida con el genotipo de interés, es decir, incrementar la persistencia de la pradera mediante la estabilidad en la densidad de tallos (Rojas et al., 2016), misma que, depende de la respuesta de los procesos de formación, muerte y supervivencia de estos (Ramírez et al., 2020).
En este sentido, la frecuencia e intensidad de corte son variables que generan cambios en la composición y permanencia de la pradera (Cruz-Hernández et al., 2017), ya que, conforme existe mayor densidad de tallos, se incrementa el rendimiento de forraje y mantiene una estabilidad poblacional de tallos (Morales et al., 2012). La investigación tuvo como objetivo determinar el efecto de la intensidad de corte en la dinámica poblacional de tallos de tres híbridos de Urochloa (Mulato II, Cayman y Mavuno) en condiciones de temporal.
El estudio se realizó de marzo de 2020 a febrero de 2021 en la Posta Zootécnica ‘Ingeniero Herminio García González’, ubicada en el municipio de Güémez, Tamaulipas, México (23° 56’ 17.55” latitud norte, 99° 06’ 2.45” longitud oeste), a 167 msnm. El clima del lugar es de tipo semiárido [BS1 (h’) hw con lluvias en verano y hasta 10% en invierno (Vargas et al., 2007)].
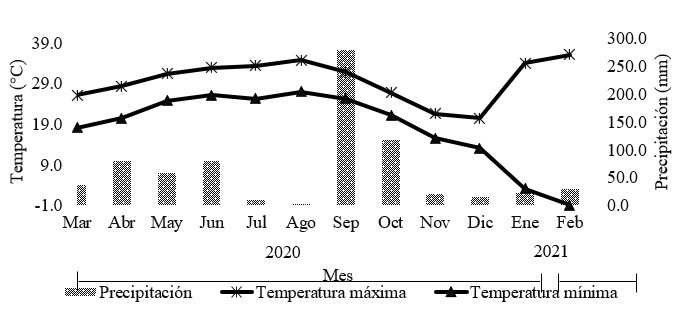
El suelo es de textura arcillosa (11.3, 23.3 y 65.4% de arena, limo y arcilla, respectivamente), con pH de 8.3, relación de adsorción de sodio es de 0.19, 4.2% de materia orgánica, 0.25% de N y 7.4, 288.6, 1.4 y 0.5 mg kg-1 de P, K, Fe y Zn, respectivamente (Garay-Martínez et al., 2018). La temperatura mínima promedio oscila entre 10 y 26 °C, la máxima entre 18 y 34 °C y la precipitación promedio acumulada del lugar es de 750 mm anuales (Figura 1).
Las praderas de Urochloa híbrido Mulato II, Cayman y Mavuno establecidas, fueron sembradas en hileras espaciadas a 0.30 m y de forma continua dentro de la hilera, a una densidad de siembra de 5 kg ha-1. Durante el experimento no se realizaron aplicaciones de fertilización o riego. Al inicio de la investigación, durante el mes de marzo de 2020, se realizó un corte para uniformizar las praderas y definir las áreas de los tratamientos según los niveles de intensidad, 15 y 25 cm, posteriormente, las fechas de conteo de tallos se realizaron cada 4 semanas y se realizaron cortes de homogenización según la intensidad definida. Los seis tratamientos definidos por los tres cultivares y dos intensidades de corte, se distribuyeron en 18 parcelas experimentales de 3 x 3 m con tres repeticiones.
Después de realizar el corte de uniformidad (marzo, 2020), en cada unidad experimental se colocaron dos cuadros fijos de alambre de 10 x 10 cm y se marcaron los tallos vivos presentes en el área con anillos del mismo color, definiéndolos como población inicial y cada cuatro semanas (fechas de muestreo) los hijuelos se marcaron con un nuevo color, conformado cada fecha una generación de tallos, mientras que, los tallos muertos se eliminaron del área de muestreo y se registró el dato.
Los datos obtenidos se utilizaron para calcular los cambios en la densidad poblacional de tallos (DPT; tallos m2), tasa de ahijamiento (TA) y muerte de tallos (TM) entre cada fecha de evaluación con las siguientes fórmulas de acuerdo con las fórmulas descritas por Ramírez et al. (2020):
La tasa de sobrevivencia de tallos (TST) se obtuvo de la siguiente manera:
El índice de estabilidad de la población de tallos (IEP) se obtuvo con la siguiente fórmula:
Donde: el IEP es la relación entre la población final o actual y la población anterior de tallos observada, considerando la tasa de supervivencia de tallos y tasa de ahijamiento. Su valor indica el efecto del ahijamiento y muerte de los tallos con relación a la densidad total, para un periodo determinado, valores próximos o iguales a 1 (0.95 a 1.05), indican una población de tallos en equilibrio y una estabilidad negativa y positiva presenta valores menores a 0.95 y mayores a 1.05, respectivamente.
Las variables de respuesta (DPT, TA, TM, TST e IEP) se analizaron usando PROC GLM (SAS, 2002) mediante un análisis de varianza con un diseño en bloques completos al azar y arreglo factorial (3 × 2 × 12) donde los factores de estudio fueron los cultivares (Mulato II, Cayman y Mavuno), las intensidades de corte (15 y 25 cm) y las fechas de muestreo (12 fechas), además, se realizó la prueba de Tukey (p= 0.05) para la comparación de medias.
En los híbridos de Urochloa se encontraron diferencias entre intensidades y fechas de muestreo para la variable densidad de tallos, tasa de ahijamiento, e índice de estabilidad (p< 0.05), mientras que, en la tasa de muerte y tasa de sobrevivencia únicamente se presentaron diferencias entre las intensidades del híbrido Mavuno, con diferencias entre fechas de muestreo en los seis tratamientos (p< 0.05).
La densidad poblacional de tallos promedio fue mayor en la intensidad de 15 cm en 11.4, 4.7 y 11.0% en los híbridos Mulato II, Cayman y Mavuno, respectivamente, en comparación a la intensidad de 25 cm (Cuadro 1). Además, se presentó una disminución al final de la evaluación (febrero-2021) respecto a la población inicial (marzo-2020) con 31 y 51% en la intensidad de 15 y 25 cm, respectivamente, en Mulato II, 39 y 75% para Cayman y 23 y 32% para Mavuno en las mismas intensidades (Cuadro 1).
| Tratamientos | |||||||||||||
| Muestreo | Mulato II | Cayman | Mavuno | ||||||||||
| 15 cm | 25 cm | 15 cm | 25 cm | 15 cm | 25 cm | ||||||||
| Mar. 20 | 2 683 | ab | 2 833 | a | 3 433 | a | 3 983.0 | a | 2 558 | a | 2 646 | a | |
| Abr. 20 | 2 483 | ab | 2 650 | ab | 3 500 | a | 3 733.0 | ab | 2 050 | bc | 1 975 | ab | |
| May. 20 | 2 133 | ab | 2 533 | ab | 2 800 | abcd | 2 967.0 | cd | 1 816 | c | 1 408 | b | |
| Jun. 20 | 2 250 | ab | 2 400 | ab | 2 994 | abc | 3 100.0 | bcd | 1 991 | bc | 1 537 | ab | |
| Jul. 20 | 2 811 | a | 2 250 | ab | 2 694 | bcd | 2 800.0 | cd | 2 083 | abc | 1 750 | ab | |
| Ago. 20 | 2 717 | a | 2 433 | ab | 2 850 | abcd | 3 033.0 | cd | 2 408 | ab | 2 150 | ab | |
| Sep. 20 | 2 639 | ab | 2 050 | abc | 2 616 | cd | 3 133.0 | bcd | 2 258 | abc | 2 500 | ab | |
| Oct. 20 | 2 955 | a | 2 350 | ab | 2 483 | cd | 3 166.0 | bc | 2 291 | abc | 2 287 | ab | |
| Nov. 20 | 2 911 | a | 2 227 | ab | 2 566 | cd | 3 033.0 | cd | 2 158 | abc | 2 050 | ab | |
| Dic. 20 | 2 733 | a | 2 033 | abc | 2 700 | bcd | 2 483.0 | e | 2 442 | ab | 1 991 | ab | |
| Ene. 21 | 2 383 | ab | 1 916 | bc | 2 383 | cd | 1 833.0 | e | 2 158 | abc | 1 425 | b | |
| Feb. 21 | 1 847 | b | 1 375 | c | 2 094 | d | 983.0 | f | 1 966 | bc | 1 787 | ab | |
| Promedio | 2 545 | A | 2 254 | B | 2 996 | A | 2 855.6 | B | 2 200 | A | 1 959 | B | |
| EEM | 67.36 | 87.12 | 86.34 | 152.18 | 52.39 | 90.10 | |||||||
La evaluación a través del tiempo con intervalo de corte establecido definió el comportamiento en la densidad de tallos en los tres híbridos, el híbrido Mulato II mostró diferencias entre las intensidades a partir del mes de octubre, manteniéndose superior la intensidad de 15 cm en 20% en promedio hasta el mes de enero, Figura 2, por lo que, probablemente esté en función de las condiciones ambientales presentes en los meses de enero y febrero, Figura 1. Al respecto, el comportamiento de la densidad poblacional de tallos con intervalo de corte prolongado en el cultivar Mulato II en conjunto con cuatro cultivares de Urochloa, presentaron densidades superiores a los 4 000 tallos m2; no obstante, entre cultivares no se encontraron diferencias significativas (Lucio et al., 2023), por ende, para este experimento la intensidad de corte es crucial para determinar el efecto que se tiene sobre la densidad poblacional de tallos, ya que se ha establecido que después de efectuarse una defoliación mecánica o mediante pastoreo, se induce al crecimiento de la densidad de tallos, lo cual, transcurre hasta los 21 y 28 días en Andropogon gayanus por ejemplo (Ramírez et al., 2020).
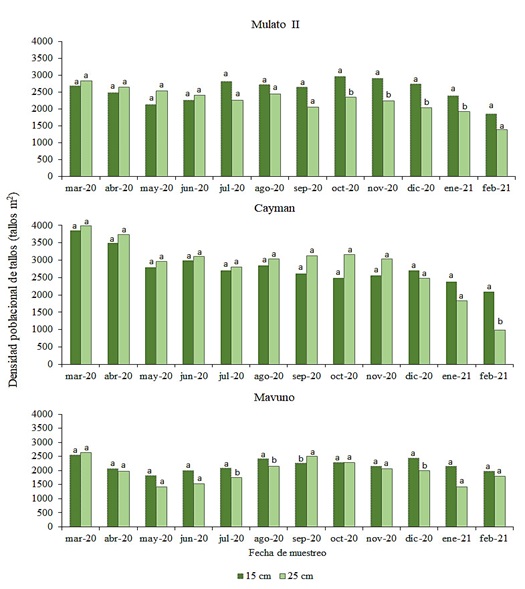
En lo que respecta al híbrido Cayman, no presentó diferencias entre intensidades (p≤ 0.05) durante la evaluación, a excepción del mes de febrero, en la cual, la intensidad de 15 cm fue superior 53% comparado con la intensidad de 25 cm, mismo comportamiento se presentó en otro estudio al evaluar el cultivar Cayman en el periodo de junio a diciembre con intensidad fija de 15 cm e intervalo de cuatro semanas (Lucio-Ruiz et al., 2021), igual que en este experimento no se tuvo diferencias entre estas fechas, por lo que se plantea que la intensidad de corte para este cultivar es de 15 cm sin afectar la densidad poblacional de tallos a través de todo el año, así mismo, esta intensidad ha sido relacionada con la obtención de rendimientos de hasta 8.8 t ha-1 de materia seca acumulada durante la época de máxima precipitación en Güémez, Tamaulipas (Garay-Martínez et al., 2018).
El híbrido Mavuno, presentó variaciones en cuanto densidad poblacional de tallos en los meses de julio, agosto y diciembre favoreciendo a la intensidad de 15 cm, mientras que, en el mes de septiembre la intensidad de 25 cm fue superior 11%, Figura 2, sugiere una respuesta a la precipitación acumulada en el mes de septiembre, dado que al tener menor densidad por área en el muestreo anterior y al propiciarse las condiciones favorables, se incrementa la densidad de tallos, misma respuesta se presentó en el pasto Mulato (36061), al ser evaluados en la época de nortes, seca y lluvias, la mayor densidad de tallos estuvo presente en la época de lluvias independientemente de la intensidad de corte (Cruz-Hernández et al., 2017).
Al evaluar el híbrido Urochloa Mulato a intensidades de 10 y 14 cm de altura, Cruz-Hernández et al. (2017) encontraron que la intensidad de 10 cm genera mayor densidad de tallos (entre 3 280 y 5 729 tallos m2); sin embargo, la precipitación en este lugar se presentó en mayor cantidad durante los meses de junio a octubre con más de 1 400 mm, mientras que, en el lugar de este estudio existe un promedio de precipitación anual de 750 mm, aunado a las temperaturas extremas, por ende, debido a las condiciones ideales para la emergencia de hijuelos, genera una mayor densidad de tallos mediante la etapa de rebrote (Ramírez et al., 2011). Durante esta etapa, sucede la activación de la zona meristemática activa ubicada en la base de los tallos, gracias a la presencia de los carbohidratos de reserva (Rincón et al., 2008).
Las mayores tasas de ahijamiento se presentaron en la intensidad de 15 cm en promedio, en comparación a la de 25 cm, además, durante el mes de septiembre se presentó las mayores tasas de ahijamiento en ambas intensidades y para los tres híbridos, debido a que es el mes de máxima acumulación de precipitación, lo cual, favorece al ahijamiento de nuevos tallos (Cuadro 2).
| Tratamientos | |||||||||||||
| Muestreo | Mulato II | Cayman | Mavuno | ||||||||||
| 15 cm | 25 cm | 15 cm | 25 cm | 15 cm | 25 cm | ||||||||
| Tasa de ahijamiento | |||||||||||||
| Abr. 20 | 2.1 | d | 2.1 | b | 8.1 | bc | 5.2 | bc | 20.2 | ab | 3.9 | d | |
| May. 20 | 18.1 | bcd | 5.3 | ab | 10.7 | bc | 8.1 | bc | 9.1 | bc | 12.2 | bc | |
| Jun. 20 | 23.2 | bc | 10.1 | ab | 20.9 | ab | 11.3 | b | 14.4 | abc | 9.1 | bcd | |
| Jul. 20 | 27.0 | ab | 11.7 | ab | 14.0 | abc | 3.2 | c | 16.5 | abc | 8.8 | bcd | |
| Ago. 20 | 17.5 | bcd | 12.7 | ab | 23.7 | a | 22.3 | a | 21.9 | ab | 29.1 | a | |
| Sep. 20 | 42. | a | 15.8 | ab | 25.0 | a | 24.7 | a | 29.0 | a | 29.6 | a | |
| Oct. 20 | 31.1 | ab | 4.8 | ab | 4.8 | c | 7.5 | bc | 25.4 | a | 14.0 | b | |
| Nov. 20 | 9.4 | cd | 5.1 | ab | 8.1 | c | 2.7 | c | 18.3 | abc | 5.6 | cd | |
| Dic. 20 | 8.5 | cd | 7.5 | ab | 9.5 | bc | 2.3 | c | 15.6 | abc | 6.3 | cd | |
| Ene. 21 | 3.2 | d | 7.8 | ab | 2.0 | c | 3.7 | c | 4.5 | c | 11.0 | bc | |
| Feb. 21 | 11.0 | cd | 27.5 | a | 4.8 | c | 4.8 | bc | 23.9 | ab | 3.2 | d | |
| Promedio | 17.6 | A | 10.0 | B | 12.0 | A | 8.7 | B | 18 | A | 11.1 | B | |
| Eem | 2.2 | 1.6 | 1.4 | 1.3 | 1.6 | 1.9 | |||||||
| Tasa de muerte | |||||||||||||
| Abr. 20 | 7.9 | bc | 8.8 | cde | 6.3 | bcd | 12.5 | cde | 24.7 | abcd | 28.9 | ab | |
| May.20 | 0.0 | c | 0.0 | e | 20.4 | abcd | 19.3 | cd | 10.4 | cde | 6.9 | bc | |
| Jun. 20 | 18.3 | b | 19.0 | c | 22.6 | abc | 7.0 | e | 0.5 | e | 0.0 | c | |
| Jul. 20 | 18.7 | b | 18.3 | cd | 23.3 | abc | 12.4 | cde | 25.0 | abc | 10.3 | bc | |
| Ago. 20 | 4.5 | bc | 8.5 | de | 20.2 | abcd | 6.4 | e | 5.5 | de | 8.1 | bc | |
| Sep. 20 | 52.5 | a | 30.5 | b | 25.2 | abc | 19.1 | cd | 31.8 | ab | 17.4 | abc | |
| Oct. 20 | 15.4 | bc | 7.1 | e | 9.6 | bcd | 8.5 | de | 5.7 | cde | 13.1 | abc | |
| Nov. 20 | 11.0 | bc | 5.3 | e | 4.7 | cd | 5.0 | e | 15.8 | bcde | 21.0 | abc | |
| Dic. 20 | 3.8 | bc | 1.2 | e | 1.2 | d | 21.9 | bc | 1.3 | e | 7.9 | bc | |
| Ene. 21 | 21.6 | b | 47.2 | a | 26.0 | ab | 31.5 | b | 18.3 | bcde | 38.0 | a | |
| Feb. 21 | 41.1 | a | 45.4 | a | 39.5 | a | 51.6 | a | 41.7 | a | 37.8 | a | |
| Promedio | 17.7 | A | 17.4 | A | 18.4 | A | 17.8 | A | 16.4 | A | 17.8 | A | |
| EEM | 2.8 | 2.8 | 2.2 | 2.4 | 2.85 | 3.0 | |||||||
En este sentido, durante el mes de agosto, donde se presenta el fenómeno conocido como canícula, acompañado de temperaturas elevadas, las cuales provocan mayor acumulación térmica para los pastos (Lucio et al., 2023) y la ausencia de precipitaciones (Pacheco-Hernández et al., 2020), los híbridos Cayman y Mavuno presentaron la mayor tasa de ahijamiento en ambas intensidades (p< 0.05), posiblemente por la utilización de las reservas de carbohidratos presentes en la base de los tallos, corona y raíces de las plantas, como una medida de perpetuar (Rincón et al., 2008). Caso contrario sucedió con el cultivar híbrido Cobra, el cual, a medida que hubo restricción de humedad y aumento de temperaturas, la tasa de ahijamiento disminuyó 12% en promedio (Cámara-Acosta et al., 2022).
La tasa de muerte presentó variaciones a través de las fechas de muestreo (p< 0.05) y fue el mes de febrero, el que presentó la mayor tasa de muerte en todos los cultivares e intensidades de corte. Cuadro 2, como resultado de las temperaturas mínimas presentes anteriores a la fecha de muestreo, Figura 1, asimismo, generó menor sobrevivencia con valores menores al 65% en los seis tratamientos en la misma fecha. En el efecto de la intensidad por cada cultivar, la intensidad de 25 cm provocó disminución en el promedio de 3% respecto a la intensidad de 15 cm solamente en la sobrevivencia de tallos del híbrido Mavuno, por lo que, exponer a este cultivar a intensidad de 25 cm repercutirá en la sobrevivencia de la población de tallos de una fecha de corte a otra (Cuadro 3).
| Tratamientos | |||||||||||||
| Muestreo | Mulato II | Cayman | Mavuno | ||||||||||
| 15 cm | 25 cm | 15 cm | 25 cm | 15 cm | 25 cm | ||||||||
| Tasa de sobrevivencia | |||||||||||||
| Abr. 20 | 91.7 | abc | 91.1 | ab | 91.4 | ab | 88.0 | ab | 75.25 | bcde | 71.1 | d | |
| May. 20 | 100.0 | a | 100.0 | a | 85.3 | bcd | 89.8 | ab | 89.5 | abc | 90.9 | abc | |
| Jun. 20 | 81.6 | bc | 82.8 | b | 80.2 | cde | 93.3 | ab | 99.4 | a | 99.5 | a | |
| Jul. 20 | 83.8 | bc | 81.6 | b | 76.6 | de | 87.5 | ab | 74.9 | cde | 91.0 | abc | |
| Ago. 20 | 95.4 | ab | 91.5 | ab | 89.7 | ab | 93.5 | a | 94.4 | ab | 91.8 | ab | |
| Sep. 20 | 54.0 | d | 69.4 | c | 74.7 | e | 80.8 | ab | 68.1 | de | 82.5 | bcd | |
| Oct. 20 | 87.6 | abc | 92.8 | ab | 87.3 | abc | 88.6 | ab | 94.2 | abc | 89.0 | abc | |
| Nov. 20 | 88.9 | abc | 91.3 | ab | 95.2 | a | 93.0 | ab | 85.9 | abcd | 78.9 | cd | |
| Dic. 20 | 96.1 | ab | 98.7 | a | 92.4 | ab | 79.5 | bc | 97.8 | a | 92.0 | ab | |
| Ene. 21 | 80.5 | c | 63.9 | cd | 73.9 | e | 66.3 | cd | 81.6 | abcd | 56.7 | e | |
| Promedio | 83.9 | A | 83.6 | A | 82.8 | A | 83.1 | A | 83.6 | A | 80.6 | B | |
| EEM | 2.4 | 2.4 | 1.7 | 2.2 | 2.8 | 3.6 | |||||||
| Índice de estabilidad | |||||||||||||
| Abr. 20 | 0.91 | bcd | 0.94 | ab | 1.00 | dc | 0.90 | ab | 0.90 | bcd | 0.74 | d | |
| May. 20 | 1.16 | a | 1.04 | a | 0.91 | cde | 0.90 | ab | 0.95 | abc | 1.02 | ab | |
| Jun. 20 | 1.02 | abc | 0.88 | abc | 0.91 | cd | 1.05 | a | 1.14 | a | 1.09 | ab | |
| Jul. 20 | 1.00 | abcd | 0.90 | ab | 0.84 | def | 0.90 | ab | 0.83 | cd | 1.00 | bc | |
| Ago. 20 | 1.15 | a | 1.00 | a | 1.14 | a | 1.06 | a | 1.10 | ab | 1.21 | a | |
| Sep. 20 | 0.69 | e | 0.79 | bcd | 0.93 | cd | 0.94 | ab | 0.88 | cd | 1.04 | ab | |
| Oct. 20 | 1.12 | ab | 0.99 | a | 0.93 | cd | 0.96 | ab | 1.13 | a | 1.01 | bc | |
| Nov. 20 | 1.00 | abcd | 0.97 | a | 1.01 | bc | 0.98 | a | 1.01 | abc | 0.81 | cd | |
| Dic. 20 | 1.03 | abc | 0.97 | a | 1.05 | ab | 0.80 | bc | 1.12 | ab | 1.01 | bc | |
| Ene. 21 | 0.86 | cde | 0.73 | cd | 0.74 | f | 0.72 | cd | 0.84 | cd | 0.63 | de | |
| Feb. 21 | 0.79 | de | 0.63 | d | 0.79 | fe | 0.61 | d | 0.72 | d | 0.45 | e | |
| Promedio | 0.98 | A | 0.89 | B | 0.93 | A | 0.89 | B | 0.96 | A | 0.91 | B | |
| EEM | 0.02 | 0.02 | 0.02 | 0.02 | 0.03 | 0.04 | |||||||
Asimismo, se ha reportado que la intensidad de 5 cm en el cultivar Mavuno tiene efectos negativos en la densidad de población total, por lo tanto, se refleja en menor tasa de sobrevivencia y este comportamiento genera cambios en la composición morfológica, ya que a intensidades de 5 cm genera tallos de mayor peso, mientras que, a intensidades de 15 cm el peso por tallo disminuye, no obstante, en el peso total de materia seca, no existen diferencias entre intensidades (Rodrigues et al., 2021). En este sentido, Cámara-Acosta et al. (2022) mencionan que para lograr la persistencia de una pradera es necesario un equilibrio entre la tasa de ahijamiento y la sobrevivencia de los tallos para mantener la densidad de tallos en determinadas situaciones.
Por otro lado, otras fechas difieren en el comportamiento de la tasa de muerte en cada cultivar por intensidad de corte, dicho comportamiento se atribuye a la competencia por espacio, a la poca o nula disponibilidad de humedad y por estrés a exposición de temperaturas inferiores a 15 °C, determinada como la temperatura base de los pastos tropicales (Lucio et al., 2023). Además, se ha encontrado que, en cortes con intervalos fijos se tiene una mayor presión sobre la tasa de muerte dado que se debilitan más los tallos (Cámara-Acosta et al., 2022).
El índice de estabilidad permite establecer una relación entre los procesos de supervivencia y la tasa de ahijamiento, ya que por sí misma no permite aclarar si es suficiente para remplazar la tasa de muerte y mantener la densidad poblacional de tallos (Ramírez et al., 2011). Por lo tanto, la intensidad de corte tuvo efecto sobre la estabilidad de tallos (p< 0.05) y la intensidad de 15 cm fue la que presentó mayor índice de estabilidad con 9, 4 y 5% respecto a la intensidad de 25 cm en los tres cultivares (Cuadro 3).
No obstante, el híbrido Cayman presentó valores menores a los 0.95; es decir, en el periodo de evaluación se obtuvo una estabilidad negativa en la pradera, por lo que, las intensidades para este híbrido no favorecen la permanencia de la pradera; mientras que, los híbridos Mulato II y Mavuno se mantuvieron en equilibrio con valores de 0.98 y 0.96. Al respecto, Lucio-Ruiz et al. (2021) al evaluar dos intensidades de corte definidas por una intensidad fija de 15 cm y una al 50% considerando la altura antes del corte en el híbrido Cayman, esta última intensidad propició mayor estabilidad de la población de tallos, ya que, las intensidades fueron menores a 15 cm de residuo, por ende, se establece que para este cultivar en el ambiente establecido se requiere mayor intensidad para incrementar el índice de estabilidad.
En el mes de agosto, los seis tratamientos presentaron una estabilidad de tallos positiva, a pesar de ser el mes donde se presenta una escasa o nula precipitación y las temperaturas son máximas, debido a la relación existente entre la mayor tasa de ahijamiento en comparación a la tasa de muerte en esta fecha (Cuadro 2). Al evaluar Panicum maximum cv. Mombaza, Ramírez et al. (2011) encontraron mayor estabilidad durante la época de sequía que en lluvias, mientras que, al evaluar dos alturas de corte, Difante et al. (2008) encontraron que la altura de 15 cm la población de tallos se mantiene más estable, con una disminución en la variación en la densidad poblacional de tallos cuando las condiciones son limitantes para la estabilidad.
El índice de estabilidad mostró los contrastes existentes entre las condiciones climáticas, dado que con déficit hídrico y temperaturas elevadas (octubre) se presentó una menor actividad en la tasa de muerte y un aumento en la tasa de ahijamiento, mientras que, durante el mes de septiembre, con las condiciones ideales para realizar recambio de tejido, se produjo una mayor tasa de muerte respecto a la tasa de ahijamiento, posiblemente por el crecimiento de las plantas y la competencia existente entre los recursos disponibles como radiación, suelo y agua (Rueda et al., 2018).
La intensidad de corte de 15 cm permitió mantener la estabilidad de la población de tallos de los híbridos Urochloa Mulato II y Mavuno. La densidad de tallos fue diferente entre fechas de muestreo según las condiciones ambientales y la densidad presente en el área, dado que, cuando se presentan las condiciones favorables de precipitación y temperatura, se incrementa la tasa de muerte, mientras que, a condiciones restringidas se produce un aumento en la aparición de hijuelos.
Cámara-Acosta, J.; Enríquez-Quiroz, J. F.; Rueda-Barrientos. J. A.; Ortega-Jiménez, E.; Ramírez-Bribiesca, J. E. y Guerrero-Rodríguez, J. de D. 2022. Dinámica de tallos del pasto Urochloa híbrido cv. cobra en respuesta al tiempo de corte. Rev. Fitotec. Mex. 45(3):303-311.
J. Cámara-Acosta J. F. Enríquez-Quiroz J. A. Rueda-Barrientos E. Ortega-Jiménez J. E. Ramírez-Bribiesca J. de D Guerrero-Rodríguez 2022Dinámica de tallos del pasto Urochloa híbrido cv. cobra en respuesta al tiempo de corteRev. Fitotec. Mex453303311
Carmona-Muñoz, M. I.; Trejo-López, C.; Ramírez-Vallejo, G.; García-de los Santos, G. y Becerril-Pérez, C. .2003. Resistencia a sequía de Brachiaria spp. I. Aspectos fisiológicos. Rev. Fitotec. Mex. 26(3):153-159.
M. I. Carmona-Muñoz C. Trejo-López G. Ramírez-Vallejo G. García-de los Santos C Becerril-Pérez 2003Resistencia a sequía de Brachiaria spp. I. Aspectos fisiológicosRev. Fitotec. Mex263153159
Cruz-Hernández, A. A.; Hernández-Garay, H.; Vaquera-Huerta, A.; Chay-Canul, J. F.; Enríquez-Quiroz, J. y Ramírez-Vera. S. 2017. Componentes morfogenéticos y acumulación del pasto mulato a diferente frecuencia e intensidad de pastoreo. Rev.Mex. Cienc. Pec. 8(1):101-109. https://doi.org/10.22319/rmcp.v8i1.4310.
A. A. Cruz-Hernández H. Hernández-Garay A. Vaquera-Huerta J. F. Chay-Canul J. Enríquez-Quiroz S Ramírez-Vera 2017Componentes morfogenéticos y acumulación del pasto mulato a diferente frecuencia e intensidad de pastoreoRev.Mex. Cienc. Pec8110110910.22319/rmcp.v8i1.4310
Cuartas, C. C. A.; Naranjo, J. F. R.; Tarazona, A. M. M. y Barahona, R. R. 2014. Uso de la energía en bovinos pastoreando sistemas silvopastoriles intensivos con Leucaena leucocephala y su relación con el desempeño animal. Rev. CES Medicina Veterinaria y Zootecnia. 8(1):70-81.
C. C. A. Cuartas J. F. R. Naranjo A. M. M. Tarazona R. R Barahona 2014Uso de la energía en bovinos pastoreando sistemas silvopastoriles intensivos con Leucaena leucocephala y su relación con el desempeño animalRev. CES Medicina Veterinaria y Zootecnia817081
Difante, G. S.; do Nascimento Junior, D.; Silva, S. C. da; Euclides, V. P. B.; Zanine, A. de M. and Adese, B. 2008. Dinâmica do perfilhamento do capim-marandu cultivado em duas alturas e três intervalos de corte. Rev. Bras. Zootec. 37(2):189-196. https://doi.org/10.1590/S1516-35982008000200003.
G. S. Difante D. do Nascimento Junior S. C. da Silva V. P. B. Euclides A. de M. Zanine B Adese 2008Dinâmica do perfilhamento do capim-marandu cultivado em duas alturas e três intervalos de corteRev. Bras. Zootec37218919610.1590/S1516-35982008000200003
Garay-Martínez, J. R.; Joaquín-Cancino, S.; Estrada-Drouaillet, B.; Martínez-González, J. C.; Joaquín-Torres, B. M.; Limas-Martínez, A. G. y Hernández-Meléndez, J. 2018. Acumulación de forraje de pasto Buffel e híbridos de Urochloa a diferente edad de rebrote. Ecosis. Rec. Agropec. 5(15):573-581. https://doi.org/10.19136/era.a5n15.1634.
J. R. Garay-Martínez S. Joaquín-Cancino B. Estrada-Drouaillet J. C. Martínez-González B. M. Joaquín-Torres A. G. Limas-Martínez J Hernández-Meléndez 2018Acumulación de forraje de pasto Buffel e híbridos de Urochloa a diferente edad de rebroteEcosis. Rec. Agropec51557358110.19136/era.a5n15.1634
Lucio, R. F.; Joaquín, C. S.; Garay, M. J. R.; Bautista, M. Y.; Estrada, D. B. y Limas, M. A. G. 2023. Dinámica de tallos e índice de estabilidad en cinco cultivares de Urochloa en condiciones semiáridas. Trop. Grassl. -Forrajes Trop. 11(1):1-10. https://doi.org/10.17138/TGFT(11)1-10.
R. F. Lucio C. S. Joaquín M. J. R. Garay M. Y. Bautista D. B. Estrada M. A. G Limas 2023Dinámica de tallos e índice de estabilidad en cinco cultivares de Urochloa en condiciones semiáridasTrop. Grassl. -Forrajes Trop11111010.17138/TGFT(11)1-10
Lucio-Ruiz, F.; Garay-Martínez, J. R.; Bautista-Martínez, Y.; Estrada-Drouaillet, B.; Hernández-Guzmán, F. J; Limas-Martínez, A. G. y Joaquín-Cancino, S. 2021. Estabilidad en la población de tallos en cultivares de Urochloa con diferente intensidad de corte. Ecosis. Rec. Agropec. Número Especial II: e2960. https://doi.org/10.19136/era.a8nII.2960.
F. Lucio-Ruiz J. R. Garay-Martínez Y. Bautista-Martínez B. Estrada-Drouaillet F. J Hernández-Guzmán A. G. Limas-Martínez S Joaquín-Cancino 2021Estabilidad en la población de tallos en cultivares de Urochloa con diferente intensidad de corteEcosis. Rec. Agropec. Número Especial IIe296010.19136/era.a8nII.2960
Morales, N. C. R.; Melgoza, A. C.; Jurado, P. G.; Martínez, M. S. y Avendaño, C. A. 2012. Caracterización fenotípica y molecular de poblaciones de zacate punta blanca (Digitaria californica (Benth.) Henr.). Rev. Mexic. Cienc. Pec. 3(2):171-184.
N. C. R. Morales A. C. Melgoza P. G. Jurado M. S. Martínez C. A Avendaño 2012Caracterización fenotípica y molecular de poblaciones de zacate punta blanca (Digitaria californica (Benth.) Henr.)Rev. Mexic. Cienc. Pec32171184
Pacheco-Hernández, A.; Garay-Martínez, J. R.; Granados-Rivera, L. D.; Bautista-Martínez, Y.; Joaquin-Torres, B. M..; Limas-Martínez, A. G. y Joaquín, S. C. 2020. Dinámica de ahijamiento de Megathyrsus maximus cv. Tanzania cosechado a diferente edad de rebrote. Ciencia e innovación. 3(1):23-31.
A. Pacheco-Hernández J. R. Garay-Martínez L. D. Granados-Rivera Y. Bautista-Martínez B. M.. Joaquin-Torres A. G. Limas-Martínez S. C Joaquín 2020Dinámica de ahijamiento de Megathyrsus maximus cv. Tanzania cosechado a diferente edad de rebroteCiencia e innovación312331
Quero, C. A. R.; Enríquez, J. F. y Miranda, L. 2007. Evaluación de especies forrajeras en América tropical, avances o estatus quo. Interciencia. 32(8):566-571.
C. A. R. Quero J. F. Enríquez L Miranda 2007Evaluación de especies forrajeras en América tropical, avances o estatus quoInterciencia328566571
Ramírez, R. O.; Silva, S. C. da; Hernández, G. A..; Enríquez, Q. F. J.; Perez, P. J.; Quero, C. A. R. y Herrera, H. J. G. 2011. Rebrote y estabilidad de la población de tallos en el pasto Panicum maximun CV. “Mombaza” cosechado en diferentes intervalos de corte. Rev. Fitotec. Mex. 34(3):213-2020.
R. O. Ramírez S. C. da Silva G. A.. Hernández Q. F. J. Enríquez P. J. Perez C. A. R. Quero H. J. G Herrera 2011Rebrote y estabilidad de la población de tallos en el pasto Panicum maximun CV. “Mombaza” cosechado en diferentes intervalos de corteRev. Fitotec. Mex3432132020
Ramírez, R. O.; Flores, I. A.; Hernández, E. C.; Rojas, A. R. G.; Maldonado, M. Á. P. y Valenzuela, J. L. L. 2020. Dinámica poblacional de tallos e índice de estabilidad del pasto llanero. Rev. Mex. Cienc. Agríc. 11(24):23-34. https://doi.org/10.29312/remexca.v0i24.2355.
R. O. Ramírez I. A. Flores E. C. Hernández A. R. G. Rojas M. Á. P. Maldonado J. L. L Valenzuela 2020Dinámica poblacional de tallos e índice de estabilidad del pasto llaneroRev. Mex. Cienc. Agríc1124233410.29312/remexca.v0i24.2355
Rincón, C. A.; Ligarreto, M. G. A. y Garay, E. 2008. Producción de forraje en ellos pastos Brachiaria decumbens cv. Amargo Brachiaria brizantha cv. Toledo, sometidos a tres frecuencias y a dos intensidades de defoliación en condiciones del piedemonte llanero colombiano. Rev. Facultad Nacional de Agronomía Medellín. 61(1):4336-4346.
C. A. Rincón M. G. A. Ligarreto E Garay 2008Producción de forraje en ellos pastos Brachiaria decumbens cv. Amargo Brachiaria brizantha cv. Toledo, sometidos a tres frecuencias y a dos intensidades de defoliación en condiciones del piedemonte llanero colombianoRev. Facultad Nacional de Agronomía Medellín61143364346
Rodrigues, L. F.; Vendramini, J. M. B.; Santos, A. C.; Dubeux Jr, J. C. B.; Miotto, F. R. C.; Sousa, L. F. and Alencar, N. M. 2021. Canopy characteristics of Mavuno hybrid brachiariagrass and Marandu palisadegrass harvested at different harvest intensities. Trop. Grassl.-Forrajes Trop. 9(3):249-255. https://doi.org/10.17138/TGFT(9)249-255.
L. F. Rodrigues J. M. B. Vendramini A. C. Santos J. C. B. Dubeux Jr F. R. C. Miotto L. F. Sousa N. M Alencar 2021Canopy characteristics of Mavuno hybrid brachiariagrass and Marandu palisadegrass harvested at different harvest intensitiesTrop. Grassl.-Forrajes Trop9324925510.17138/TGFT(9)249-255
Rojas, G. A. R.; Hernández, G. A.; Quero, C. A. R.; Guerrero, R. J. D.; Zaragoza, R. J. L. y Trejo, L. C. P. 2016. Persistencia de Dactylis glomerata L. solo y asociado con Lolium perenne L. y Trifolium repens L. Rev. Mex. Cienc. Agríc. 7(4):885-895.
G. A. R. Rojas G. A. Hernández C. A. R. Quero R. J. D. Guerrero R. J. L. Zaragoza L. C. P Trejo 2016Persistencia de Dactylis glomerata L. solo y asociado con Lolium perenne L. y Trifolium repens LRev. Mex. Cienc. Agríc74885895
Rueda, J. A.; Ortega-Jiménez, E.; Enríquez-Quiroz, J. F.; Palacios-Torres, R. E. and Ramírez-Ordóñes. S. 2018. Tiller population dynamics in eight cultivars of elephant Grass during undisturbed growth. Afr. J. Range Forage Sci. 35(2):1-11. https://doi.org/10.2989/10220119.2018.1477832.
J. A. Rueda E. Ortega-Jiménez J. F. Enríquez-Quiroz R. E. Palacios-Torres S Ramírez-Ordóñes 2018Tiller population dynamics in eight cultivars of elephant Grass during undisturbed growthAfr. J. Range Forage Sci35211110.2989/10220119.2018.1477832
Vargas, T. V.; Hernández, R. M. E.; Gutiérrez, L. J.; Plácido, D. C. J. y Jiménez, C. A. 2007. Clasificación climática del estado de Tamaulipas, México. Ciencia UAT. 2(2):15-19.
T. V. Vargas R. M. E. Hernández L. J. Gutiérrez D. C. J. Plácido C. A Jiménez 2007Clasificación climática del estado de Tamaulipas, MéxicoCiencia UAT221519
Velázquez, M. M.; Hernández, F. J. G.; Cervantes, J. F. B. y Guillermo, H. G. V. 2015. Establecimiento de pastos nativos e introducidos en zonas semiáridas de México. Instituto Nacional de Investigaciones Forestales Agrícolas (INIFAP)-CIRNE-Campo Experimental San Luis. México. 22 p.
M. M. Velázquez F. J. G. Hernández J. F. B. Cervantes H. G. V Guillermo 2015Establecimiento de pastos nativos e introducidos en zonas semiáridas de MéxicoInstituto Nacional de Investigaciones Forestales Agrícolas (INIFAP)CIRNE-Campo Experimental San LuisMéxico22 p