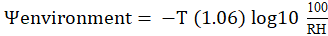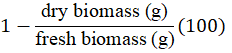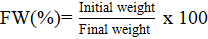Revista Mexicana Ciencias Agrícolas volume 13 number 7 September 28 - November 11, 2022
DOI: https://doi.org/10.29312/remexca.v13i7.3083
Article
Moisture content and rehydration in rose flower stems
exposed to the environment
Mayte Stefany Jiménez-Noriega1
Gumercindo Honorato De-La-Cruz-Guzmán2§
Alberto Arriaga-Frías2
Manuel Mandujano-Piña2
Alfredo León-Macedo2
1Botanical Garden-Faculty of Higher Studies Cuautitlán-UNAM. Cuautitlán-Teoloyucan Highway km 2.5, San Sebastián Xhala, Cuautitlán Izcalli, State of Mexico, Mexico. CP. 54714. (mayajimenez13@yahoo.com).
2Unit of Morphology and Function-Faculty of Higher Studies Iztacala-UNAM. Av. de los Barrios num. 1, Los Reyes Iztacala, Tlalnepantla, State of Mexico, Mexico. CP. 54090. (ixbe@unam.mx; manuelm@unam.mx; alfredoleonmacedo@gmail.com).
Corresponding author: moashi@unam.mx.
Abstract
At least in one phase of postharvest, the flower stems of rose are exposed to the environment. The water they lose and the rehydration in the vase will depend on the temperature and anatomy of the flower stem. In this research, the moisture content of two cultivars of Rosa hybrida exposed to room temperature from 0 to 48 h was evaluated and the anatomy of the leaf and stem was described. In March 2019, flower stems of ‘Engagement’ and ‘Freedom’ roses were harvested, in a commercial greenhouse in Puebla, Mexico and transported dry to the laboratory of the Iztacala Faculty of Higher Studies, where they were spread out on a table. Evaluations were conducted every 8 h. In both cultivars, the initial moisture of stems and leaves was 75%. In those that were exposed for 24 hours, it was 66 and 57% for ‘Engagement’ and ‘Freedom’, respectively. The thickness of the cuticle and mesophyll was 96 and 26% greater in ‘Engagement’ than in ‘Freedom’ (180.9 and 2.4 μm). The stems of ‘Engagement’ had 331 vessels per mm2 with 29.5 μm in diameter, those of ‘Freedom’ 402 of 26.5 μm. The ‘Engagement’ rose lost less water than ‘Freedom’ due to the greater thickness of its cuticle and amplitude of its mesophyll. The number of vessels had little effect on water loss or rehydration in the vase. In both cultivars, the flower stems could have a useful vase life even if they are exposed to the environment for 8 or 16 h without any protection, with longer times it decreases due to dehydration of the leaves.
Keywords: Rosa hybrida L., cuticle thickness, number of vessels, vase life.
Reception date: June 2022
Acceptance date: August 2022
Introduction
During postharvest management, flower stems must keep their moisture content so that their visual appearance is attractive to the consumer (Walton et al., 2010). Dehydration is avoided by wrapping the flower stems with insulating materials such as Kraft paper and black polyethylene bags or by immersing them in moisturizing solutions (Mosqueda-Lazcares et al., 2011; De La Cruz-Guzmán et al., 2018). However, it is common that, at least at some stage of the postharvest, producers or distributors expose the flower stems to the environment without any protection. Under these conditions, dehydration is accelerated, and the moisture content decreases due to the water potential gradient (ΔΨ) that is generated between the flower stems and the environment.
The water potential of the environment (Ψenvironment) is regulated by temperature and relative humidity (RH) and is calculated with the equation:
.
Where: T= absolute temperature; RH= relative humidity (Taiz and Zeiger, 2010). The water potential recorded in flower stems of ‘Freedom’ rose, freshly harvested, fluctuates between -0.7 and -0.2 MPa (Arévalo-Hernández, 2011). In case these stems are exposed to an environment with water potential of -123.6 MPa; that is, with a temperature of 20 °C and RH of 40%, the ΔΨ between the stems and the environment will have a value of -123.15 MPa. This value generates such a great force that water will flow from the inside of the flower stems into the environment and would dehydrate them in a few minutes.
To avoid dehydration, stomata closure is an almost immediate response to water stress. However, between species there are no differences in the stomatal index, in addition to the fact that, in flower stems, the closure capacity depends on the relative humidity values in their preharvest phase (Wilkinson, 1979; Fanourakis et al., 2012; Fanourakis et al., 2013). Therefore, the anatomical characteristics of the leaf and stem that do change during the crop cycle are a useful tool to explain why some flower stems hydrate faster than those of others when placed in the vase (Moreno, 2009).
For example, thicker leaf cuticle or wider mesophyll decrease water loss (Tafolla-Arellano et al., 2013). In the stem, the diameter and number of vessels is related to the velocity of water flow (Zimmermann, 1983). Thus, a group of vessel elements whose diameters are two times larger will increase the water flow up to 24 times, according to Poiseuille’s law, and will have greater participation in the rehydration of the flower stems, compared to the vessels of smaller diameter (Nijsse et al., 2000; Arriaga-Frías et al., 2016), a similar response will occur with flower stems of ‘Lovely Red’, ‘Topaz’ and ‘Rouge Baiser’ roses, which have 267, 289 and 308 vessels per mm2, respectively (Cohen et al., 2012; De La Cruz-Guzmán et al., 2016; De La Cruz-Guzmán et al., 2018).
In flower stems that are exposed to the environment without any protection, the water status will depend on the exposure times, anatomy and genetics of the cultivars, among others. Anatomical differences could lead to not all flower stems rehydrating or that they do so at different times when placed in a vase (Hernández-Hernández et al., 2009; Mosqueda-Lazcares et al., 2011; De La Cruz-Guzmán et al., 2018).
Changes in the fresh weight of flower stems after harvest have been described by different authors (Mosqueda-Lazcares et al., 2011; Arriaga-Frías et al., 2016; De La Cruz-Guzmán et al., 2018), but none of them explains the relationship between moisture content and the anatomy of the stem or leaf. Therefore, the objective of this research was to evaluate the moisture content in the postharvest of two cultivars of Rosa hybrida L. (Rosales Bercht. & J. Presl; Rosaceae Juss.) exposed to the environment, for different periods of time and its relationship with the anatomy of the flower stem.
Materials and methods
In March 2019, fifty-nine flower stems (55 to 60 cm in length) of ‘Engagement’ and ‘Freedom’ roses were harvested in a commercial greenhouse of the company Flores de la Sierra Mágica de Puebla (http://floressierramagica.com), located in the municipality of Ahuazotepec, Puebla, Mexico. Immediately, all the flower stems with full foliage were wrapped with Kraft paper and black polyethylene bags to be transported to the Iztacala Faculty of Higher Studies, of the National Autonomous University of Mexico (UNAM). In the laboratory where the experiment was carried out (temperature between 22.1 and 25.2 °C, RH of 15 to 16.5% and water potential of -259.6 to -245.3 MPa), all the flower stems or experimental units were unpacked and spread out on a table randomly.
The treatments were the times of exposure to the environment: 0 (at the time of unwrapping), 8, 16, 24, 32, 40 and 48 h. In each treatment, the following response variables were evaluated: a) moisture content (MC). For each treatment, three flower stems from each cultivar were randomly selected, fractionated into leaves, stem and flower bud and weighed with a Velab® ES-1000H digital scale, then dried in a Reavel® oven, REA-II2240 (both from Científica Velaquin, CDMX, Mexico) at 80 °C for 24 h to record the dry biomass and calculate the MC with the equation: MC(%)=
b) fresh weight and rehydration in the vase. From each treatment or evaluation time, five flower stems of ‘Engagement’ and ‘Freedom’ roses were randomly selected, cut to 50 cm in length, the lower foliage was removed to leave three pentafoliate and three trifoliate leaves, the initial weight (iw) of each stem (experimental unit) was recorded and it was placed in a bottle with 250 ml of ‘universal’ tap water for exclusive use in cut flower experiments (Van Meeteren and van Gelder, 2001) and they were randomly distributed on the tables of the laboratory. Their final weight (fw) was obtained twenty-four hours later, and the fresh weight (FW) was calculated in percentage with the equation:
The flower stems were considered as rehydrated as long as their leaves recovered their turgor without showing symptoms of wilt or that their FW was similar to 100%; c) opening index (OI, dimensionless) and vase life (VL, days). The following were obtained from the flower stems from each treatment that were placed in ‘universal’ tap water and rehydrated: their flower opening index by recording with a digital vernier (Truper®), the upper diameter of their flower buds and it was divided by the maximum flower opening of each cultivar (De La Cruz-Guzmán et al., 2015). The VL was determined by counting the number of days the stems remained in the bottles without showing symptoms of senescence such as bent-neck, petal drooping or leaf wilt; and d) leaf and stem anatomy. At the time of unwrapping the flower stems and before spreading them out on the table, three of each cultivar were selected and placed for 3 h in ‘universal’ tap water to completely restore their hydration.
In the middle part of each stem and in the middle leaflets of its pentafoliate leaves, samples were taken and fixed in FAA (formaldehyde, 10%; acetic acid, 5%; absolute alcohol, 50%), 48 h later they were washed with water and stored in 70% alcohol. On the leaves, freehand cross-sections were made, and they were stained with safranin and fast green. Stem samples were embedded in methyl acrylate (Zarlavsky, 2014; Jiménez-Noriega et al., 2019) and cross-sections of 1 μm thickness were performed with rotary microtome (Leica 2051, Westlar, Germany), staining was performed with cresyl violet. Paradermal samples were obtained by scraping the surface of the leaves and staining them with safranin.
The terminology and qualitative anatomical descriptions of the leaves were based on Metcalfe and Chalk (1980, 1983), for those of the stems, the recommendations of (IAWA Committee 1989) were followed. For the evaluations of the quantitative anatomical characteristics, 30 repetitions per individual were performed, using an image analyzer (Image-Pro Plus version 6.1, Media Cybernetics Inc., Maryland, USA). The anatomical variables of the leaf that were measured were thickness of the cuticle, length and width of adaxial and abaxial epidermal cells, width of the palisade, spongy parenchyma and of the leaf taking the cuticle of both surfaces of the leaf blade as reference. While in the stem, the thickness of the cuticle, number of vessels per mm2, area and radial or tangential diameters of the vessel elements were measured. Statistical analysis. Descriptive statistics, one-factor analysis of variance and mean comparison tests (Tukey, p≤ 0.05) were applied to the results with the SAS® 9.0 software for Windows.
Results and discussion
Moisture content
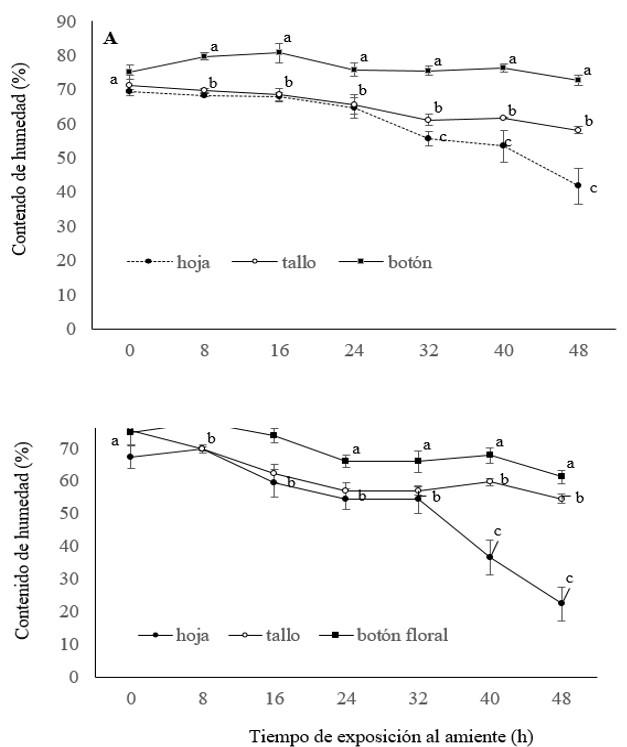
The stems of ‘Engagement’ rose of all treatments or times of exposure to the environment kept the moisture of their flower buds between 73 and 75%. In the leaves and stems, from treatments 0, 8, 16 or 24 h, the moisture decreased from 70 to 66%. For those of 24, 32, 40 and 48 h, moisture loss was greater in leaves (from 65.69 to 41.7%) than in stems (from 65.2 to 58.2%) (Figure 1A).
In ‘Freedom’ rose, the moisture of the flower buds decreased from 75.4 to 61.2% in all times of exposure to the environment. In the stems and leaves of this cultivar, moisture loss was from 74.4 to 54.4 and from 67.3 to 22.4 respectively (Figure 1B). In both cultivars, the leaves were the organs that lost the most moisture and the flower buds those that kept it the longest. It is worth mentioning that the flower buds, leaves and stems of ‘Freedom’ rose lost more moisture compared to those of ‘Engagement’ (Figure 1A and 1B).
De la Cruz-Guzmán et al. (2018) mention that the morphology of the flower buds and the compact arrangement of the petals allows them to maintain moisture for longer, compared to stems or leaves which have a larger surface area exposed to the environment and are therefore more susceptible to dehydration. At all times of exposure to the environment (0 to 48 h), dehydration was higher in the leaves of ‘Freedom’ rose (67.3 to 22.4%) than in ‘Engagement’ (from 69.5 to 41.7%).
Figure 1. Moisture content in leaves, stems and flower buds of ‘Engagement’ (A) and ‘Freedom’ (B) roses exposed to the environment from 0 to 48 h. Means with different letters, in each evaluation time, indicate significant differences (Tukey, p≤ 0.05). Each data represents the average of three repetitions ± standard error.
Because both cultivars were exposed to the same water potential gradient (≈ -252 MPa), the lower moisture loss in ‘Engagement’ can be explained because its cuticle was two times bigger than that of ‘Freedom’ (Table 1). The cuticle alone acts as a barrier to the loss of water to the atmosphere, the thicker it is, the less foliar dehydration and the tissues will keep their moisture content for longer (Sperry, 2003; Tafolla-Arellano et al., 2013). In the leaves of Rosa hybrida, epicuticular waxes that, together with the cuticle, increase the resistance to water loss and maintain the integrity of the cell membranes have been described (Taiz and Zeiger, 2010).
Wax content can vary from cultivar to cultivar. For example, in ‘Lovely Red’ rose they are more abundant than in ‘Rouge Baiser’ rose (Cohen et al., 2012). The above suggests that ‘Engagement’ rose, in addition to having thicker cuticle, could have a higher proportion of epicuticular waxes than ‘Freedom’. The width of the leaf of ‘Engagement’ rose was 26% greater than in ‘Freedom’ (180.9 μm), this because the thickness of the cuticle, length of the adaxial, abaxial epidermal cells and the width of the palisade parenchyma were 49, 41, 45.3 and 18.3% greater in ‘Engagement’ rose than in ‘Freedom’ (Table 1). These anatomical differences contributed to the longer retention of moisture in ‘Engagement’ rose, particularly because the wider mesophyll allowed it to store more water and its cuticle, thicker, lose it slowly compared to ‘Freedom’.
Table 1. Quantitative anatomy of the leaf in two cultivars of Rosa hybrida.
Rosa hybrida (cultivars) | Cuticle (μm) | Adaxial E. (μm) | Abaxial E. (μm) | Parenchyma (μm) | Leaf (μm) | |||||
Length | Width | Length | Width | Palisade | Spongy | |||||
Engagement | 4.7 az | 39.1 a | 28.5 a | 34 a | 27.8 a | 79.8 a | 57.5 a | 243.6 a | ||
Freedom | 2.4 b | 23.1 b | 19.5 b | 18.6 b | 14.5 b | 65.2 b | 56.5 a | 180.9 b | ||
HSD | 0.5 | 2.6 | 2.6 | 2.9 | 2.8 | 3.1 | 4.4 | 5.1 | ||
CV (%) | 30.1 | 27.6 | 22.3 | 37.8 | 45.1 | 14.7 | 26.1 | 8.1 | ||
z= means with different letters, in each column, indicate significant differences (Tukey, p≤ 0.05). E= epidermis; HSD= honest significant difference; CV= coefficient of variation.
The reduction in leaf width indicates that the plants developed in xeromorphic environments or that they were exposed to a period of water stress during their cultivation (Ely et al. 2005; Moreno, 2009). In contrast, thicker leaves are typical of well-irrigated plants, they have greater capacity to store water and lose it slowly if their cuticle is thicker (Moreno, 2009). This could suggest that the flower stems of ‘Freedom’ rose remained in water stress at some stage of the preharvest, while those of ‘Engagement’ rose were well irrigated or had shorter periods of stress. Differences in anatomy are also attributed to the genetic characteristics of each cultivar, so the above statement would be valid for wild species and be used with greater caution in cultivars whose purpose is commercialization, since the producer will always seek to provide the best preharvest management.
Description of leaf anatomy
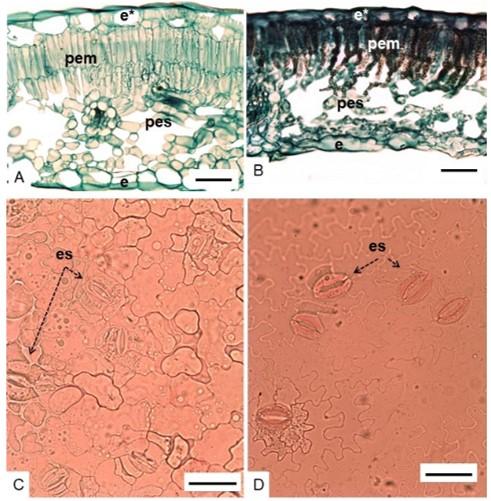
Both cultivars of Rosa hybrida had smooth cuticle, unistratified epidermis (Figure 2A, 2B), paracytic-type stomata, with two cells annexed to the same level as the epidermal ones (Metcalfe and Chalk, 1979) (Figure 2C, 2D), similar to what was reported by Hernández-Hernández (2009) for ‘Vega’ and ‘Grand Gala’ roses. In ‘Engagement’ and ‘Freedom’ roses the distribution was hypostomatic, as in ‘Lovely Red’ and ‘Rouge Baiser’ roses (Cohen et al., 2012).
Figure 2. Anatomy of the leaf of ‘Engagement’ (A, C) and ‘Freedom’ (B, D) roses. Cross section (A, B); paradermal view (C, D); e*= adaxial epidermis; e= abaxial epidermis; pem= palisade parenchyma; pes= spongy parenchyma; es= stomata; scale= 50 μm.
The location of the stomata on the abaxial surface of the leaves contributes to the decrease in the transpiratory rate since they do not directly receive sunlight (Willmer and Fricket, 1996; Raven, 2002). Dark contents, possibly tannins, were observed on the leaves of ‘Freedom’ rose, which were not visible in ‘Engagement’.
Fresh weight (FW) and rehydration in the vase
In treatments with 8 or 16 h of exposure to the environment, the flower stems of ‘Engagement’ rose had an average FW of 118.5%, in that of 24 h, the FW increased to 138.6%, which can be attributed to the dehydration of the xylem. Until the 24 h treatment, all the flower stems rehydrated when placed in the vase. In those of 32, 40 or 48 hours, the fresh weight was 107.8, 99.6, 97.8% and the percentage of stems that rehydrated was 60, 40 and 0, respectively. For ‘Freedom’ rose, in treatments 8, 16 or 24 h of exposure to the environment, the FW of the flower stems fluctuated from 112.6 to 128.3% and 100% of the stems rehydrated. With those of 40 or 48 h, the FW decreased to 96% and no flower stem rehydrated when placed in the vase (Table 2).
Table 2. Fresh weight and rehydration of flower stems of ‘Engagement’ and ‘Freedom’ roses exposed to the environment for 48 h.
Time (h) | ‘Engagement’ | ‘Freedom’ | |||
Fresh weight (%) | Rehydration (%) | Fresh weight (%) | Rehydration (%) | ||
8 | 119.5 b | 100 | 112.6 ab | 100 | |
16 | 117.5 bc | 100 | 115.1 ab | 100 | |
24 | 138.6 a | 100 | 128.3 a | 100 | |
32 | 107.8 bcd | 60 | 102.1 b | 20 | |
40 | 99.6 cd | 40 | 96 b | 0 | |
48 | 97.8 d | 0 | 96 b | 0 | |
HSD | 18.6 | - | 20.6 | - | |
CV (%) | 8.4 | - | 9.5 | - | |
z= means with different letters, in each column, indicate significant differences (Tukey, p≤ 0.05). HSD= honest significant difference; CV= coefficient of variation. Each value represents the average of five repetitions.
The increase in fresh weight depends on the absorption rate that occurs in response to the water potential gradient (ΔΨ) between the vase solution and the xylem of the stem (Taiz and Zeiger, 2010; Van Doorn, 2012). Exposure to the environment dehydrates the flower stems and decreases the moisture content of the xylem, which contributes to faster absorption when placed in the vase. ‘Engagement’ and ‘Freedom’ roses were exposed to the same environment at the laboratory table, so their dehydration was regulated by the thickness of the cuticle, width of the mesophyll and the number, area or diameter of the vessel elements.
The number of vessels per mm2 was lower in the stem of ‘Engagement’ rose (331) than in that of ‘Freedom’ (402). In addition, the cuticle thickness, area and diameters of the vessel elements were 10.8, 16.6 and 9.7% higher in ‘Engagement’ than in ‘Freedom’, whose cuticle had a thickness of 9.1 μm, while the area and diameter of the vessel elements corresponded to 520.9 mm2 and 25.2 μm, respectively (Table 3).
Table 3. Quantitative anatomy in the middle part of the stem of two cultivars of Rosa hybrida.
Cultivars of Rosa hybrida | Cuticle thickness (μm) | Vessel elements | Vessel diameter (μm) | |||
Number mm-2 | Area (μm2) | Radial | Tangential | |||
Engagement | 10.2 az | 330.7 b | 624.2 a | 26.3 a | 29.5 a | |
Freedom | 9.1 b | 401.6 a | 520.9 b | 23.9 b | 26.5 b | |
HSD | 0.7 | 27.1 | 46.6 | 1.9 | 2.2 | |
CV (%) | 14.9 | 14.2 | 26.1 | 26.1 | 26.8 | |
z= means with different letters, in each column, indicate significant differences (Tukey, α≤ 0.05). HSD= honest significant difference; CV= coefficient of variation.
Short vessel elements with larger diameters are more efficient in conducting water, since shorter lengths give resistance to the collapse of the cell wall, by preventing its deformation (Carlquist, 1977; Sperry, 2003).
In contrast, the reduction in the diameter of the vessel elements in ‘Freedom’ rose (Table 3) indicates a lower water conductivity, but ensures the flow of water, because they are less susceptible to cavitate (Hargrave et al., 1994; Nijsse et al., 2000; Martínez-Vilalta and Piñol, 2002). The number and diameter of the vessel elements contribute directly to the hydration of the flower stems, since vessel elements with diameters greater than 75 μm have lower resistance to water flow (van Door and Reid, 1995; Van Meeteren, 1999). The diameter of the vessels of ‘Engagement’ or ‘Freedom’ roses were within the range that Hernández-Hernández et al. (2009) and Cohen et al. (2012) report for ‘Grand Gala’, ‘Vega’, ‘Lovely Red’ and ‘Rouge Baiser’ roses, which range between 13 and 67.7m.
Description of the anatomy of the stem
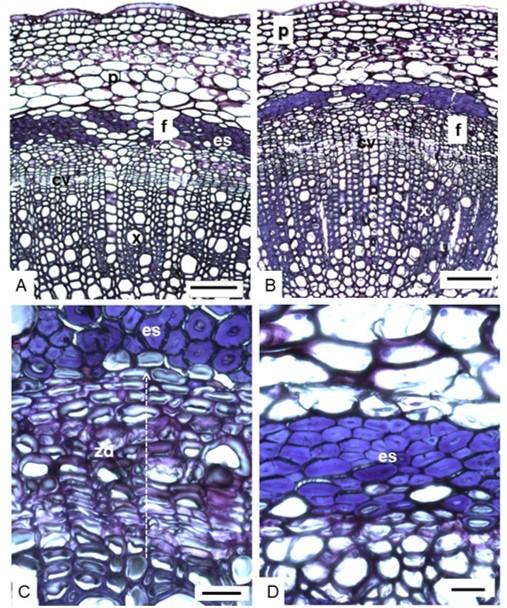
Both cultivars of Rosa hybrida had thick cuticle and unistratified epidermis. The parenchyma of the cortex is interrupted by packages of seven strata of sclereid cells in cell differentiation, in ‘Engagement’ rose and four strata without differentiation in ‘Freedom’ (Figure 3C, 3D). Below the sclereids was the secondary phloem divided by rays from the pith. In xylem, porosity was diffuse (Figure 3A, 3B).
The differentiation of tissues, which involves the development of cells to the thickening of the secondary walls, is related to vascular cambium activity and this with the radial and longitudinal growth of the stem (Jiménez-Noriega et al., 2019), which suggests that the flower stems of ‘Engagement’ rose at the time of harvest were growing and therefore with higher metabolic activity than ‘Freedom’, which could also promote the absorption of water after exposing them to the environment and placing them in the vase.
Figure 3. Anatomy of the stem of ‘Engagement’ (A, C) and ‘Freedom’ (B, D) roses. Cross section (A, B) 100 μm; zone of differentiation of the sclereids (C, D) 20 μm. e= epidermis; p= parenchyma; es= sclereids; f= phloem; cv= vascular cambium; zd= zone in differentiation; x= xylem.
Flower opening index (OI) and vase life (VL)
The flower stems of ‘Engagement’ rose, which rehydrated after 8 or 16 hours of being exposed to the environment, had a flower opening index of 0.68 ±0.21 and a vase life of 10.2 d, while the OI and VL of the flower stems of ‘Freedom’ rose were 0.78 ±0.15 and 8.2 ±1.96 d, respectively. The reduction in vase life of ‘Freedom’ was attributed to differences in stem and leaf anatomy. In this work, ‘Engagement’ rose had greater vase life and the diameters of its vessels were larger compared to ‘Freedom’.
This contrasts with the results obtained by Hernández-Hernández et al. (2009), since the ‘Vega’ cultivar that had the largest diameters exhibited on average 2.6 days less vase life than ‘Grand Gala’ and they attributed it to water stress due to its susceptibility to dehydration.
OI values less than 1.0 indicate that the flower buds were below their maximum opening and greater than or equal to 1.0 that the treatment promoted their flower opening. Without applying any treatment or postharvest management, the maximum flower opening of ‘Engagement’ and ‘Freedom’ roses is 7.9 ±1.2 and 6.7 ±0.5 cm, with a vase life of 8.9 ±1.07 and 7.6 ±1.06 d (De La Cruz-Guzmán et al., 2015). This indicates that exposure to the environment for 8 or 16 h reduces the opening of the flower bud but increases the vase life of ‘Engagement’ and ‘Freedom’ roses by 1.3 and 0.6 d, respectively. Longer times of exposure to the environment reduce vase life, for example, in the 24 h treatment, all the flower stems were considered as rehydrated because the flower buds kept or restored their moisture content (Figure 1, Table 2). However, their vase life was less than or equal to 3 d for ‘Engagement’ rose and less than or equal to 2.5 d for ‘Freedom’ because the leaves dehydrated and withered more quickly.
Conclusions
When exposed to the environment without any protection, the flower stems of ‘Engagement’ rose keep their moisture content longer because their leaves have thicker cuticle and wider mesophyll than those of ‘Freedom’. In both cultivars, all the flower stems rehydrated after exposure to the environment for up to 24 h. With longer times, the number of rehydrated stems and vase life decrease due to dehydration of the leaves. In ‘Engagement’ rose, the vessel elements with diameters larger than those of ‘Freedom’ contributed to the fact that even with 32 or 40 h of exposure to the environment, there is rehydration of flower stems. The flower buds, due to their compact shape, are the organs that lose the least amount of water, while, due to their extended shape, the leaves lose the greatest amount of moisture. Due to the high water potential gradient that is generated between the environment and the flower stems of ‘Engagement’ or ‘Freedom’ roses, producers or distributors are recommended to avoid exposing them to the environment without any protection.
Acknowledgements
This work was supported by the Morphology and Function Unit of the Iztacala Faculty of Higher Studies, National Autonomous University of Mexico (UNAM), FES-Iztacala.
Cited literature
Arévalo, H. J. J. 2011. Evaluación del efecto de la aplicación de diferentes láminas de riego en el cultivo de rosa (Rosa sp.) cv Freedom, bajo invernadero en la sabana de Bogotá. Tesis de Maestría en Ingeniería Agrícola. Universidad Nacional de Colombia. Colombia. 67-71 pp. https://docplayer.es/44421466-Evaluacion-del-efecto-de-la-aplicacion-de-diferentes-laminas-de-riego-en-el-cultivo-de-rosa.html.
Arriaga, F. A.; De La Cruz, G. G. H. y Mandujano, P. M. 2016. Conductividad hidráulica en tallos florales de rosa cv ’Polo’ con diferente longitud. Rev. Fitotec. Mex. 39(3):233-239. http://www.redalyc.org/articulo.oa?id=61046936007.
Carlquist, S. 1977. Ecological factors in wood evolution: a floristic approach. Am. J. Bot. 64(7):887-896. https://www.jstor.org/stable/2442382?seq=1.
Cohen, G.; Mascarini, L. y Xifreda, C. C. 2012. Anatomía y micromorfología de hojas y tallos de dos cultivares de Rosa hybrida L. para flor de corte. Phyton. 81(2):199-204. http://www. scielo.org.ar/pdf/phyton/v81n2/v81n2a10.pdf.
De La Cruz, G. G. H.; Arévalo, G. M. L.; Peña, V. C. B.; Castillo, G. A. M.; Colinas, L. M. T. y Mandujano, P. M. 2015. Influencia del índice de cosecha en la vida de florero de siete cultivares de Rosa hybrida. Agroproductividad. 8(2):3-11. https://biblat.unam.mx/ hevila/Agroproductividad/2015/vol8/no2/1.pdf.
De La Cruz, G. G. H.; Arévalo, G. M. L.; Peña, V. C. B.; Castillo, G. A. M.; Colinas, L. M. T. y Mandujano, P. M. 2016. Influencia del manejo seco a dos temperaturas y longitud del tallo en la vida de florero de rosa ‘Polo’. Bio Ciencias. 4(1):27-39. Doi: https://doi.org/10. 15741/revbio.04.01.03.
De La Cruz, G. G. H.; Saucedo, G. D. M.; Arriaga, F. A. y Mandujano, P. M. 2018. Rehidratación y longevidad en tallos florales de rosa ‘Topaz’ y crisantemo ‘Hartman’ almacenados en seco a temperatura ambiente. Bio Ciencias. 5:1-14. http://revistabiociencias.uan.mx/ index.php/BIOCIENCIAS/article/view/316/pdf.
Ely, F.; Torres, F. y Gaviria, J. 2005. Relación entre la morfoanatomía foliar de tres especies de Miconia (Melastomataceae) con su hábitat y distribución altitudinal en el parque nacional Sierra Nevada de Mérida, Venezuela. Acta Botánica Venezuelica. 28(2):275-299. https://www.redalyc.org/articulo.oa?id=86228207.
Fanourakis, D.; Pieruschkaa, R.; Savvides, A.; Macnish, A. J.; SarlikiotI, V. and Woltering, E. J. 2013. Sources of vase life variation in cut roses: a review. Postharvest Biol. Technol. Doi: https://doi.org/10.1016/j.postharvbio.2012.12.001.
Fanourakis, D.; Carvalho, S. M. P.; Almeida, D. P. F.; Van Kooten, O.; Van Doorn, W. G. and Heuvelink, E. 2012. Posthar vest water relations in cut rose cultivars with contrasting sensitivity to high relative air humidity during growth. Postharvest Biol. Technol. 64(1):64-73. Doi: https://doi.org/10.1016/j.postharvbio.2011.09.016.
Hargrave, K. R.; Kollb, K. J.; Ewers, F. W. and Davies, S. D. 1994. Conduit diameter and drought-induce embolism in Salvia mellifera Greene (Labiatae). New Phytol. 126(4):695-705. https://nph.onlinelibrary.wiley.com/doi/10.1111/j.1469-8137.1994.tb02964.x.
Hernández, H. F.; Arévalo, G. M. L.; Colinas, L. M.T.; Zavaleta, M. H. A. y Valdés, C. J. 2009. Diferencias anatómicas y uso de soluciones de pulso en dos cultivares de rosa (Rosa sp.). Rev. Chapingo, Ser. Hortic. 15(2):11-16. https://www.scielo.org.mx/pdf/rcsh/v15nspe/ v15nspea3.pdf.
Jiménez, N. M. S.; López, M. L.; Aguilar, R. S. y Terrazas, T. 2019. Actividad cambial y cribo-xilogénesis de tres especies en un gradiente altitudinal en la Sierra Nevada, México. Bot. Sci. 97(4):725-740. Doi: https://doi.org/10.17129/botsci.2336.
Martínez, V. J. and Piñol, J. 2002. Drought-induced mortality and hydraulic architecture in pines populations of the NE Iberian Peninsula. Forest Ecol. Manag. 161(1-3):247-256. Doi: https://doi.org/10.1016/S0378-1127(01)00495-9.
Metcalfe, C.R. and Chalk, L. 1979. Anatomy of dicotyledons, systematic anatomy of leaf and stem, with a brief history of the subject. 2nd (Ed.). Clarendon Press. Oxford, Great Britain. 166-180 pp.
Metcalfe, C. R. and Chalk, L. 1980. Anatomy of the dicotyledons: systematic anatomy of leaf and stem, with a brief history of the subject. 2nd (Ed.). Clarendon Press. Oxford, Great Britain. 66-89 pp.
Metcalfe, C. R. and Chalk, L. 1983. Anatomy of the dicotyledons: wood structure and conclusion of the general introduction. 2nd (Ed.). Clarendon Press. Oxford, Great Britain. 180-201 pp.
Moreno, P. L. 2009. Respuesta de las plantas al estrés por déficit hídrico. Una revisión. Agronomía Colombiana. 27(2):179-191. https://www.redalyc.org/articulo.oa?id=180316234006.
Mosqueda, L. G.; Arévalo, G. L.; Valdovinos, P. G.; Rodríguez, P. J. E. y Colinas, León, M. T. 2011. Época de corte y manejo postcosecha de ocho cultivares de rosa de corte. Rev. Mex. Cienc. Agríc. 3(3):591-602. http://www.scielo.org.mx/pdf/remexca/v2nspe3/vspen3a15.pdf.
Nijsse, J. X.; Van Meeteren, U. and Keijzer, C. J. 2000. Air in xylem vessels of cut flowers. Acta Hortic. 517:497-486. Doi: https://doi.org/10.17660/ActaHortic.2000.517.61.
Raven, J. 2002. Selection pressures on stomatal evolution. New Phytolgist. 153(3):371-386. Doi: https://doi.org/10.1046/j.0028-646X.2001.00334.x.
Sperry, J. S. 2003. Evolution of water transport and xylem structure. Inter. J. Plant Sci. 164(3):115-127. https://sperry.biology.utah.edu/publications/IJPS%2003.pdf.
Taiz, L. and Zeiger, E. 2010. Plant physiology. 5th (Ed). Sinauer associates. Massachusetts, USA. 33-46 pp.
Tafolla, A. J. C.; González, L. A.; Tiznado, H. M. E.; Zacarías, G. L. y Báez, S. R. 2013. Composición, fisiología y biosíntesis de la cutícula en plantas. Rev. Fitotec. Mex. 36(1):3-12. http://www.scielo.org.mx/pdf/rfm/v36n1/v36n1a1.pdf.
Van Meeteren, U. and Van Gelder, A. 1999. Effect of time since harvest and handling conditions on rehydration ability of cut chrysanthemum flowers. Postharvest Biol. Technol. 16(2):169-177. Doi: https://doi.org/10.1016/S0925-5214(99)00020-4.
Van Meeteren, U. and Van Gelder, A. 2001. Should we reconsider the use of deionized water as control vase solutions? Acta Hortic. 543:257-264. https://core.ac.uk/download/pdf/ 29305998.pdf.
Van Doorn, W. G. 2012. Water relations of cut flowers: an update. Horticultural reviews. 40:55-106. Doi: https://doi.org/10.1002/9781118351871.ch2.
Walton, E. F.; Boldingh, H. L.; McLaren, G. F.; Williams, M. H. and Jackman, R. 2010. The dynamics of starch and sugar utilization in cut peony (Paeonia lactiflora Pall.) stems during storage and vase life. Postharvest Biol. Technol. 58(2):142-146. https://www.sciencedirect. com/science/article/abs/pii/S0925521410001225.
Wilkinson, H. P. 1979. The plant super face (mainly leaf). Metcalfe and Chalk (Ed.). Anatomy of dicotyledons. Oxford, Claredon Press. London. 97-165 pp.
Willmer, C. M. y Fricket, M. 1996. Stomata. 2nd (Ed,). Chapman & Hall. London, UK. 126-191 pp.
Zarlavsky, G. E. 2014. Histología vegetal. Técnicas simples y complejas. Sociedad Argentina de Botánica. Buenos Aires, Argentina. 198 p.
Zimmermann, M. H. 1983. Xylem structure and the ascent of sap. Springer-Verlag. New York, USA. 4-36 pp.