elocation-id: e2979
The use of rootstocks in modern viticulture is a common practice to avoid the impact of phylloxera and nematodes on vineyards. However, the use of rootstocks affects the metabolic processes of the plant, including the assimilation of water and ions from the soil. In Mexico, as in other grape-growing countries, the use of rootstocks has become widespread to protect vineyards against harmful microorganisms. Nevertheless, little is known about the impact of rootstock use on the physiology of vine varieties. Therefore, this study aimed to evaluate the effect of rootstocks 110-R, 101-14, 1103-P, and 3309-C on the assimilation of nitrogen, phosphorus, sodium, copper, iron, manganese, and zinc in Merlot, Shiraz, and Cabernet Sauvignon vines from Chihuahua, Mexico. The nitrogen concentration did not vary in relation to the rootstock used or the variety grown. In addition, the concentration of N in vine leaves decreased over time. This is the result of the mobilization of ions to storage areas during the period of foliar senescence. On the other hand, the concentration of phosphorus, sodium, copper, iron, and zinc in the leaves of the varieties and rootstocks studied did not present significant differences. Nonetheless, the concentration of Mn in Merlot leaves was 30% higher in vines with rootstock 1103-P compared to vines with rootstock 3309-C. In this study, it was shown that rootstock impacts ion assimilation in wine grape varieties.
Vitis vinifera L., nutrient absorption, nutrition, rootstock.
The use of rootstocks in viticulture became widespread worldwide as a response to the presence of organisms that impact the growth and survival of Vitis vinifera, including phylloxera and nematodes. At present, the use of rootstocks in viticulture responds not only to protection against predatory organisms or parasites but also to protect the vineyards against adverse soil and moisture conditions. Therefore, knowledge of soil type, nutrient concentration, moisture, viticultural practices, and insects, among others, is critical for the selection of the most suitable rootstocks in vineyards.
In general, rootstocks used in viticulture belong to the American species Vitis riparia, V. rupestris, V. berlandieri, and V. champinii (Winkler et al., 1974). In addition, there are several rootstocks that are the product of crosses between these species or between American species and Vitis vinifera (Muñoz and González, 1999). Some of these rootstocks have been very successful in protecting V. vinifera from problems of phylloxera, nematodes, acidic soils, high soil salinity, etc. (Ollat et al., 2014; Zhou-Tsang et al., 2021).
However, in Mexico, the impact of rootstocks on the assimilation of soil ions and their accumulation in plants is unknown. Ion assimilation varies substantially between American vine species and varieties of V. vinifera (Schaller and Löhnertz, 1990; Heinitz et al., 2020). Boron assimilation, for example, was up to two times lower in Pinot noir planted without rootstock in Oregon, USA, than the same variety grafted onto rootstocks Harmony, 5C, 420A, 3309C, 101-14, and 44-53 (Candolfi-Vasconcelos et al., 1997).
Similarly, the concentration of potassium in Pinot noir must was lower in vines without rootstock than in vines planted on different rootstocks at four sites in Oregon, USA (Candolfi-Vasconcelos et al., 1997). In contrast to these studies, the concentration of nitrogen, potassium, phosphorus, calcium, and magnesium in the must of Riesling grapes varied significantly in relation to the rootstock used and the planting locality (Schaller and Löhnertz, 1990).
In addition, chloride assimilation was less than half in Vitis girdiana from southern Nevada compared to rootstock 140-Ruggeri (Heinitz et al., 2020). These differences in ion assimilation are potentially linked to the differences in proton generation by the roots of each of the rootstocks, varieties, or species. Differences in the assimilation of these ions due to rootstocks on a grape variety can have a great impact on the organoleptic characteristics of the wine produced from those grapes.
In Mexico, the use of rootstocks in viticulture has become widespread as a strategy for the protection of vineyards against phylloxera and nematodes. Nevertheless, the impact of rootstocks on ion assimilation in vines and musts is unknown.
Chihuahua is a state of Mexico with a long history in the production of grapes for direct consumption and for the production of wines and distillates (Ojeda-Barrios et al., 2012). Due to the presence of phylloxera in most of the soils of Chihuahua, it has been decided to plant all vineyards with rootstocks. Nonetheless, there is not enough information on the impact of rootstocks on growth, grape ripening, water stress of vines, ion assimilation, etc.
The ions assimilated by the roots of vines planted without grafting or on rootstocks are accumulated in different tissues of plants, including fruit (Winkler et al., 1974). The accumulation of sodium in grapes from the Guadalupe Valley, Mexico, for example, has been associated with a saline perception in wines produced with this fruit (Cabello-Pasini et al., 2013).
Therefore, the analysis of the concentration of ions in tissues (leaves, petioles, fruits) is critical to determine the nutritional levels of the vines and the impact that these ions will have on the organoleptic characteristics of the wines produced with these grapes (Christensen and Kearney, 2000). Consequently, this study aimed to evaluate the effect of rootstocks 110-R, 101-14, 1103-P, and 3309-C on the assimilation of nitrogen, phosphorus, sodium, copper, iron, manganese, and zinc in Merlot, Shiraz, and Cabernet Sauvignon vines from Chihuahua, Mexico.
Samples of leaves of vine (Vitis vinifera) of the varieties Cabernet Sauvignon, Shiraz, and Merlot were collected in the vineyard of the Faculty of Agrotechnological Sciences of the Autonomous University of Chihuahua, Cuauhtémoc Campus (28° 24’ 43” north latitude, 106° 52’ 50” west longitude) during the active growth period from May to September 2018. The vines were cultivated in a double trellis structure, spaces of 1.5 m between vines and corridors of 3 m.
The experimental plants were five years old and were fertigated with a drip system. The vines were grafted onto rootstocks 110-R, 101-14, 1103-P, and 3309-C, the training was formed to two arms, and pruning was carried out in winter up to approximately 30 buds per vine.
To assess the concentration of ions in the tissue, mature leaves of the different varieties studied were collected randomly from May to September. The collected leaves were washed with deionized water to remove dust, sulfur, insecticides, and other contaminants and were dehydrated in an oven (Labline, Chicago, USA) at 60 °C to constant weight (2 d). The midrib of the leaves was removed, and the tissue was pulverized (< 1 mm) in a mill with stainless steel blades (Thomas Scientific, Minimill, USA). The material was preserved in plastic bags until analysis.
The concentration of total nitrogen in the tissue was determined by the Kjeldahl method (AOAC, 1999). Previously macerated tissue (0.1 g) was digested with 0.3 g of selenium and 5 ml of concentrated sulfuric acid for 24 h at room temperature. NaOH at 40% (w/v) was added to the samples until the mixture changed to purple, and it was immediately distilled. The vapors were collected in a 30 ml boric acid receiving solution (4% v/v) plus six drops of bromocresol green and six drops of methyl red. The total nitrogen concentration was determined by titration with HCl 0.02 N.
The total phosphorus concentration was determined by the molybdovanadate yellow method (AOAC, 1999). Samples of 1 g of tissue were digested in 25 ml of a triacid solution (H2SO4 89%, HCl 9%, HClO4 2% v/v) for 8 h at room temperature. Digested samples of 5 ml were reacted with 10 ml ammonium molybdate and 10 ml ammonium metavanadate and made up to 50 ml.
The solution was incubated for 10 min at room temperature, and absorbance was determined at 470 nm (Hash, DR 5000 UV-Vis, USA). The phosphorus concentration of the samples was determined using a monopotassium phosphate calibration curve.
The concentration of Sodium, Copper, Iron, Manganese, and Zinc in the tissue of the varieties studied was estimated by atomic absorption spectrophotometry (AOAC, 1999). To 1 g of sample, 25 ml of a triacid mixture (nitric acid 89%, perchloric acid 9%, and sulfuric acid 2% v v) was added, and it was left to stand for 8 h at room temperature. The samples were filtered (Whatman 40, cellulose, 8 μm) and made up to 50 ml with distilled water. The concentration of the ions was determined in the atomic absorption spectrophotometer (Perkin Elmer Analysis 2000, USA).
The effects of rootstock and grape variety in ion assimilation were assessed using a one-way Anova after checking the homoscedasticity (Bartlett) and normality (Kolmogorov-Smirnov) of the data. Multiple comparisons were evaluated using Tukey’s post hoc tests (Sokal and Rohlf, 1995) in SigmaStat (Systat Software, San Jose, CA, USA).
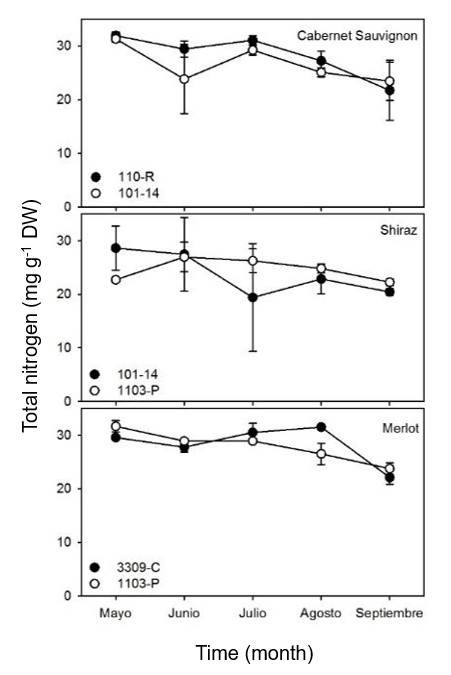
No significant differences (p> 0.05) were observed in the concentration of total nitrogen in the leaves of the three varieties studied in relation to the different rootstocks used (Figure 1). As observed in this study, the concentration of N in Malbec tissue planted without grafting or grafted onto six different rootstocks in Mendoza, Argentina, did not show significant differences either (Di Filippo, 2008).
Despite the similarity in N concentration in leaves and petioles, this variety showed differences in yield, foliar hydrostatic pressure, and water use efficiency.
The concentration of N in the leaves of the vines studied was approximately 2-3% and is consistent with that reported in other studies (Holzapfel and Treeby, 2007; Ozdemi et al., 2010). Overall, the concentration of N in Cabernet Sauvignon, Shiraz, and Merlot leaves on the different rootstocks in this study decreased throughout the season. The concentration of N in Cabernet Sauvignon leaves decreased linearly from approximately 31 mg g-1 DW (3.1% w/w) in May to approximately 23 mg g-1 DW (2.3% w/w) in September for both rootstocks used (110-R and 101-14).
The concentration of total N decreased slightly in the Shiraz grape with rootstock 101-14, while it remained constant in the same variety with rootstock 1103-P. Like Cabernet Sauvignon, the concentration of total N in the leaves decreased significantly from 30 mg g-1 DW (3% w/w) to approximately 24 mg g-1 DW (2.4% w/w) in Merlot grafted onto rootstocks 3309-C and 1103-P. It has been shown that the concentration of N in vine tissue is regulated by the dose and source of N during fertilization, as well as the phenological stage of the plant.
In this sense, the decrease in the concentration of N found in vine tissue throughout the growth period in this study is consistent with those observed in different vine varieties in Chile, Australia, and Spain (Laibarra et al., 2004; Holzapfel and Treeby, 2007; Callejas et al., 2012). These studies found that nitrogen levels were high (approx. 4%) at the beginning of the season and decreased throughout the season to values below 2%.
Similarly, a decrease in nitrogen concentration has been observed in the tissue of cherry, apple, and other crop leaves (Millard and Thomson, 1989; Muñoz et al., 1993; Fernández et al., 2014). This is related to the senescence of the leaves in deciduous fruit trees. Plants have been shown to mobilize nitrogen from leaves to trunks, stems, and roots prior to winter defoliation (Millard and Grelet, 2010).
This storage of nitrogen and other compounds allows the availability of ions during the next growth cycle. Therefore, the decrease of N in vine leaves in this study is also the result of a strategy of mobilization of reserve products at the end of the phenological cycle.
In general, no significant differences (p> 0.05) were observed in the concentration of phosphorus in the leaves of the three varieties studied in relation to the different rootstocks used (Figure 2). The concentration of P for Cabernet Sauvignon leaves showed a linear decrease of approximately 0.06 mg g-1 DW in May to end in September with an estimated value of 0.04 mg g-1 DW for both rootstocks.
The concentration of P in Shiraz increased significantly in July for both rootstocks. In the Merlot grape, the highest concentrations of P were observed during June and July, with values above 0.08 mg g-1 DW in both rootstocks. The results of this study are consistent with those observed in other analyses where phosphorus concentrations were higher in the flowering and veraison stages and decreased over time (Romero et al., 2004; Callejas et al., 2012).
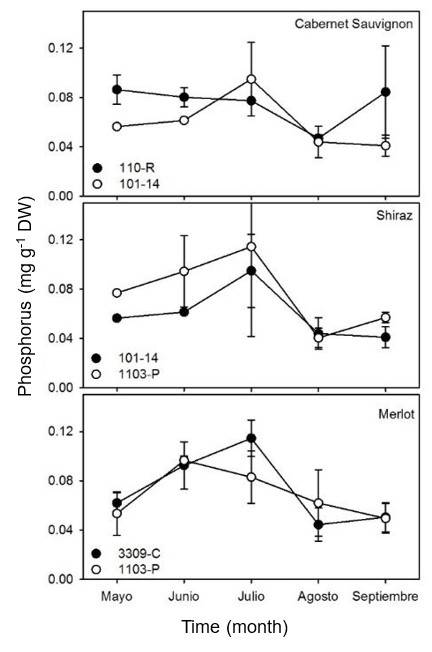
Phosphorus is an essential primary determinant of the initial growth of plant tissues, especially during the flowering stage in vines (Srinivasan and Mullins, 1981; Sierra, 2001). Therefore, it is likely that the greater assimilation of phosphorus observed during flowering in the vines of this study is the result of a physiological response to the increased demand for phosphorus during this phenological stage.
A significant increase in the concentration of Na was observed in the leaves of the three varieties studied in relation to time (Figure 3). However, there were no significant differences in the absorption of Na in relation to the different rootstocks used. For Cabernet Sauvignon leaves, the concentration of Na increased from approximately 1.5 mg g-1 DW in May to 3 mg g-1 DW in July for both rootstocks.
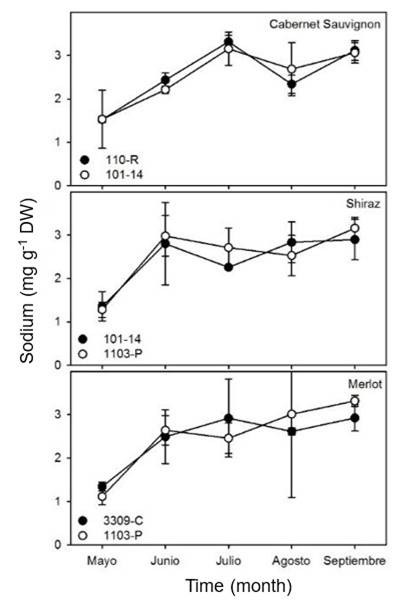
In Shiraz leaves, the lowest concentrations were observed in May, and they became saturated at approximately 3 mg g-1 DW from June to September for both rootstocks studied. The concentration of Na in Merlot was similar to that observed for Shiraz, the lowest values were observed in May, and they became saturated at approximately 3 mg g-1 DW from June. The concentration of Na found in the different varieties of vine coincides with other studies that show greater accumulations of Na from veraison (Navarro et al., 1991).
The resistance of rootstocks to high concentrations of Na varies in relation to the different genotypes, but 3309-C and 1103-P were found to be more sensitive than the rest of the rootstocks analyzed (Martin and Vila, 2013). This increase in the accumulation of Na in tissue has been shown to have an impact on photosynthetic processes and impact the growth of vines. The increased assimilation and fixation of Na in vine tissue, for example, resulted in a decrease in chlorophyll concentration in leaves (Ojeda-Barrios et al., 2012).
This reduction in chlorophyll concentration in the tissue was due to the impact on osmotic potential, which altered the physiological and biochemical functions of the plant (Fuchs et al., 2000). This suggests that the type of soil and the irrigation scheme have a greater impact on the assimilation of Na than the type of rootstock used in vine crops.
No significant differences (p> 0.05) were observed in the concentration of copper in the leaves of the three varieties studied in relation to the different rootstocks used (Figure 4). The concentration of Cu in Cabernet Sauvignon leaves remained constant over the study time at approximately 5.5 μg g-1 DW in both rootstocks. In Shiraz, Cu concentration in leaves decreased linearly from 7 μg g-1 DW to 6 mg g-1 DWin rootstocks 101-14 and 1103-P. On the other hand, the maximum Cu values in Merlot were observed in May (approx. 6 μg g-1 DW), and they decreased in September to approximately 5 μg gDW-1 in both rootstocks.
The concentrations of Cu obtained in the different vine samples are similar to those found in other studies (Romero et al., 2004; Callejas et al., 2012). These studies show that copper concentrations decrease from flowering to early veraison, with values ranging from 32 μg kg-1 to 14 μg kg-1. Cu plays catalytic roles in several important enzymes, including polyphenol oxidase and ascorbic acid oxidase (Hopkins and Hüner, 2008).
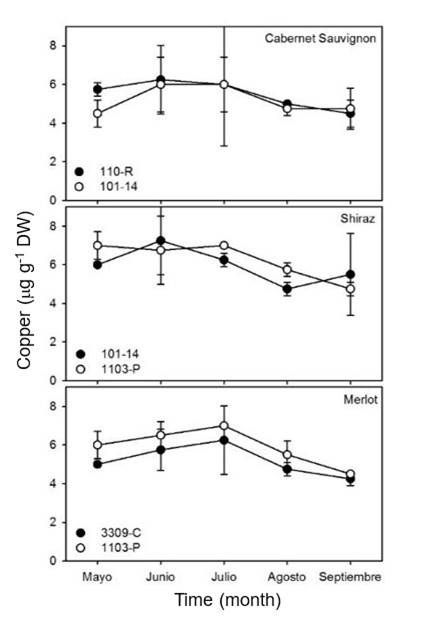
In addition, it plays a critical role in the electron transport chain during photosynthesis and may be involved in nitrite reduction (Medina et al., 1999). This suggests that Cu concentrations in the leaves of the vines studied in Chihuahua also decreased in response to the reduction of chlorophyll and senescence of the leaves.
The concentration of Fe in the leaves of the three varieties studied did not present significant variations (p> 0.05) in relation to time (Figure 5). The concentration of Fe in Cabernet Sauvignon and Shiraz leaves remained constant at approximately 250 μg g-1 DW for all rootstocks used in this study.
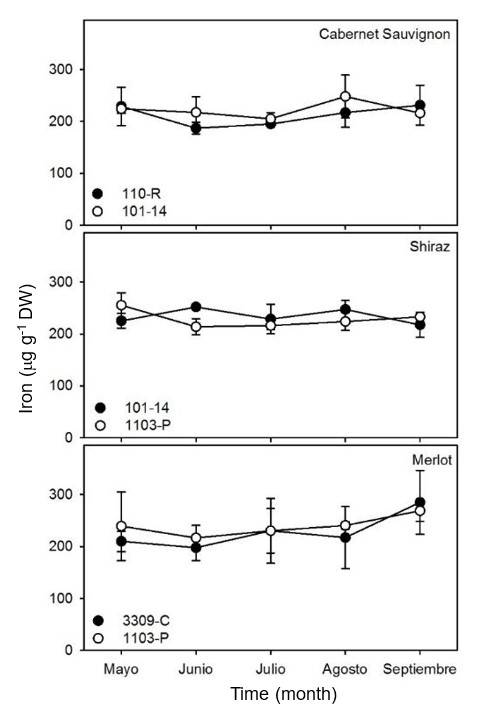
The concentrations of Fe found in the samples of vine leaves are consistent with the results observed in vines from Jerez de la Frontera in Spain (Sarmiento et al., 1992). This study shows that the concentration of Fe in vine leaves does not present a definite trend. Generally, iron deficiencies are manifested by inhibition of chlorophyll synthesis and a reduction in the rates of growth and elongation of shoots (Díaz et al., 2013). However, the concentration of Fe in soils is generally not limiting, except in areas rich in calcium carbonate.
As in other ions, no significant differences (p> 0.05) were observed in the concentration of Mn in the leaves of Cabernet Sauvignon and Shiraz with the different rootstocks used. The assimilation of Mn in Merlot was significantly higher (p< 0.05) in vines with rootstock 1103-P than in vines with rootstock 3309-C (Figure 6). For Cabernet Sauvignon leaves with rootstock 101-14, the concentration of Mn remained constant over time. Nevertheless, the concentration of Mn in Cabernet Sauvignon with rootstock 110-R increased from 30 to 60 μg g-1 DW from May to September.
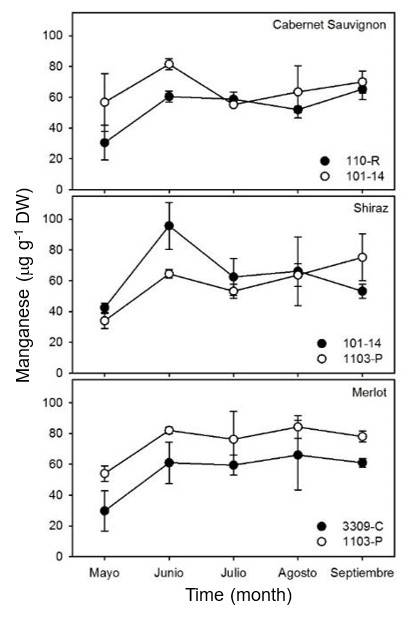
The concentration of Mn in Shiraz leaves with rootstock 101-14 increased to approximately 95 μg g-1 DW in June and decreased to 50 μg g-1 DW in September. Unlike Shiraz with rootstock 101-14, the concentration of Mn in Shiraz with rootstock 1103-P increased linearly from 30 to 60 μg g-1 DW over time. On the other hand, the concentration of Mn in Merlot leaves throughout the cycle was 30% higher (p< 0.05) in vines with rootstock 1103-P compared to vines with rootstock 3309-C. The lowest concentrations of Mn in vine leaves were observed in May, and they became saturated from June to September in Merlot leaves with both rootstocks.
The concentrations of Mn found in the samples of three vine varieties show similarity to that found in Chilean vines (Callejas et al., 2012). In general, Mn levels tend to rise in the growth stage due to the participation of Mn in the synthesis of tyrosine and its derivatives, such as indoleacetic acid (IAA) auxin, which promotes positive phototropism and the development of lateral and adventitious roots in plants.
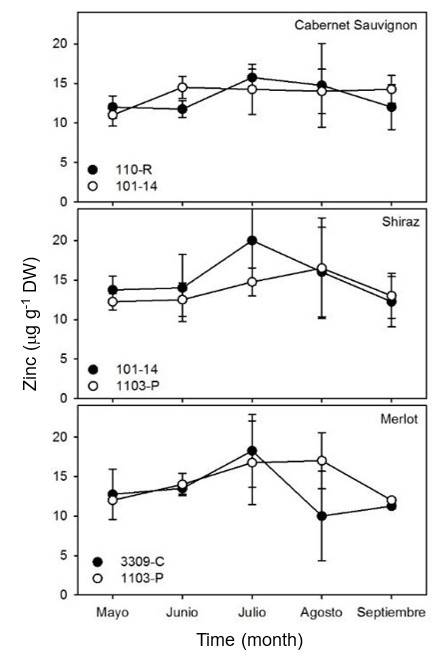
Due to the above, the availability of Mn in the roots of the plant increases in the phenological stages of fruit set and veraison (Catalina et al., 2014) and remains or decreases during fruit ripening. No significant differences (p> 0.05) were observed in the concentration of Zn in the leaves of the three varieties studied and in relation to the different rootstocks used (Figure 7). The concentration of Zn in Cabernet Sauvignon leaves in both rootstocks remained constant at approximately 17 μg g-1 DW over time.
In Shiraz, with both rootstocks studied, an increase was observed in the concentration of Zn in the leaves during July and August, and then it decreased in September. The concentration of Zn in Merlot leaves with both rootstocks remained constant over time.
The results of leaf samplings in the three different vine varieties are consistent with the data obtained in vines from La Rioja, Spain, and other regions of the world (Romero et al., 2004; Callejas et al., 2012).
These studies showed that the highest concentrations of Zn occur during fruit set (Etchevers et al., 1983; Ojeda-Barrios et al., 2012). This increase in Zn assimilation during fruit set has been observed in other crops, such as mango (Fallas et al., 2010). This suggests that the increase in the assimilation of Zn responds mainly to the phenological cycles of plants and not so much to the genotypic differences of the rootstocks analyzed in this study.
The use of rootstocks affects the metabolic processes of the plant, including the assimilation of water and ions from the soil. The study showed that rootstock can modify the absorption of some ions in some grape varieties. Specifically, manganese absorption varied significantly in the Merlot grape depending on the rootstock used.
However, the absorption of other ions did not change in the different varieties studied, regardless of the rootstock used. Clearly, the concentration of N in vine leaves decreased over time. This suggests a mobilization of ions to storage areas during the period of foliar senescence. This is the first study in Mexico of the effect of rootstocks on ion assimilation. Likewise, in addition to choosing rootstocks to protect the plant from pathogenic organisms, it is necessary to consider the effect they can have on the assimilation of ions from the soil.
Laibarra, I. R.; Rueda, I. M.; Lorenzo-Iñigo, I.; García-Oliveras, C.; Villar-Rubio, M. T.; López-Llaría, D.; Ibáñez-Pascual, S.; Arroyo-De-Grandes, M. C. y García-Escudero, D. E. 2004. Diagnóstico nutricional de “Vitis vinifera” L. CV tempranillo en la DO Ca. Rioja: niveles críticos preliminares. Zubía. 16:41-57.
Romero, L. I.; Martín, R. I.; Lorenzo, I. I.; García, O. C.; Villar, R. M. T.; López, L. D.; Ibáñez, P. S.; Arroyo, D. G. M. C. y García-Escudero, D. E. 2004. Diagnóstico nutricional de Vitis vinifera L. cv. Tempranillo en la DO Ca. Rioja: niveles críticos preliminares. Zubía Monográfico . 16:41-57.
Zhou-Tsang, A.; Wu, Y.; Henderson, S. W.; Walker, A. R.; Borneman, A. R.; Walker, R. R. and Gilliham, M. 2021. Grapevine salt tolerance. Australian J. Grape Wine Res. 27(2):149-168. https://doi.org/10.1111/ajgw.12487.