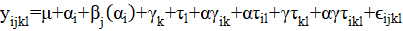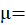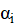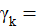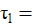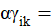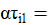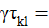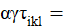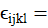Revista Mexicana de Ciencias Agrícolas volumen 13 número 4 16 de mayo - 29 de junio, 2022
DOI: https://doi.org/10.29312/remexca.v13i4.2927
10.29312/remexcav13i4.2927
Artículo
Componentes fisiológicos y numéricos del rendimiento en canola afectados
por la densidad y sistema de siembra
Gustavo García-Hernández1
Rogelio Araujo-Díaz1
Gaspar Estrada-Campuzano2§
Carlos Gustavo Martínez-Rueda2
Aurelio Domínguez-López2
1Maestría en Ciencias Agropecuarias y Recursos Naturales-Universidad Autónoma del Estado de México. (koetesfwck@hotmail.com; wardndc@hotmail.com).
2Facultad de Ciencias Agrícolas-Universidad Autónoma del Estado de México. (cgmartinezr@uaemex.mx; adominguezl@uaemex.mx).
§Autor para correspondencia: gestradac@uaemex.mx.
Resumen
En el presente trabajo se evaluaron los componentes fisiológicos y numéricos del rendimiento en canola afectados por la densidad y sistema de siembra. Se evaluaron dos genotipos primaverales de canola: Hyola 61 (híbrido) y Bioaureo 2486 (polinización abierta) bajo tres densidades 50, 75 y 90 semillas m-2), durante el ciclo invierno-primavera 2019-2020. Los tratamientos se establecieron bajo un diseño de bloques completos al azar con cuatro repeticiones, en cada uno de los dos sistemas SCP (sistema cama plana) y SCDH (sistema cama doble hilera) que fueron considerados como ambientes. El SCP presentó en promedio el mayor rendimiento (4.9 t ha-1). Los cambios en el rendimiento de semilla estuvieron asociados a una mayor producción de biomasa a madurez. Bioaureo 2486 superó en 7% el número de semillas obtenido por Hyola 61 en la densidad de 90 plantas m-2. El número de semillas por m2 estuvo asociado positivamente con el número de silicuas por m2. Sin embargo, el aumento en la densidad de plantas disminuyó el número de ramas.
Palabras clave: Brassica napus L., biomasa, densidad de población, número de semillas.
Recibido: enero de 2022
Aceptado: marzo de 202
Introducción
La canola (Brassica napus L.) es una oleaginosa reconocida por la calidad de su aceite comestible cuyo contenido de grasas esenciales puede prevenir enfermedades del corazón (Kirkegaard et al., 2016). La superficie mundial sembrada con este cultivo es de 37 579 575 ha, con una producción de más de 75 millones de toneladas y rendimiento promedio de 1.9 t ha-1 (FAOSTAT, 2018). En México, durante 2018 se cosecharon 2 058 ha, con una producción de 1 473 t y un rendimiento promedio de 0.715 t ha-1 (CONASIPRO, 2019). Los estados más sobresalientes en la producción de canola en este año fueron Tamaulipas, Estado de México, Tlaxcala, Hidalgo y Jalisco, destacando Tamaulipas con 91% de la superficie sembrada y volumen de producción; sin embargo, en esta entidad se han presentado los rendimientos más bajos (0.6 t ha-1), mientras que el Estado de México e Hidalgo obtienen los más altos (2.5 t ha-1) (CONASIPRO, 2019).
La densidad de población tiene un efecto importante sobre el crecimiento y el rendimiento de canola y depende fuertemente del arreglo espacial (distancia entre plantas y entre hileras) y del tipo de cultivar (poblaciones de polinización libre o híbridos) (Rathke et al., 2006). A nivel mundial se han evaluado distintas densidades de población en canola que van desde 8 a 190 plantas m-2, observándose un efecto diferencial entre los factores que influyen sobre el rendimiento como: la distancia entre hileras, distancia entre plantas, factores ambientales y dosis de fertilización etc.
Por ejemplo, Hosseini et al. (2006) concluyeron que un aumento en la densidad de plantas reduce significativamente la longitud de la inflorescencia principal y el número total de silicuas por planta, pero aumentó la altura de la planta, estos autores encontraron que el mayor rendimiento de semilla se obtuvo con 190 plantas m-2 y afirman que la densidad óptima de plantas aumenta la capacidad competitiva de la canola contra las malezas. De la misma manera, Mobasser et al. (2008) observaron que un incremento en la densidad de plantas disminuyó la altura de planta, diámetro del tallo, número de silicuas por planta, longitud de silicua y rendimiento de semilla. El mayor rendimiento de semilla se produjo con una densidad de 80 plantas m-2.
Del mismo modo, Kazemeini et al. (2010) determinaron que 90 plantas m-2 es la densidad óptima para el cultivo de canola, afirmando que dicha densidad influye positivamente en el peso de semilla de las silicuas secundarias que contribuye a mejorar el rendimiento de semilla y la producción de biomasa. Por otro lado, Li et al. (2014) observaron que incrementos en la densidad reducen la altura de planta, el número de ramas y silicuas por planta y el índice de cosecha; sin embargo, a mayor densidad de plantas puede mejorar la adaptabilidad de la canola al regular la estructura de la planta.
Del mismo modo, Wang et al. (2015) concluyeron que el aumento en la densidad de plantas complementado con un adecuado arreglo espacial entre parcelas e hileras puede considerarse como una práctica óptima para el incremento del rendimiento en canola. Por otro lado, existen evidencias que en el cultivo de canola es posible obtener con bajas densidades rendimientos de semilla equiparables a aquellos obtenidos con alta densidad (Gan et al., 2016; Rondanini; et al., 2017), lo cual puede ser atribuible a cambios en la morfología de planta (número de ramas). La variabilidad en los resultados obtenidos en las investigaciones previas demuestra que existe la necesidad de entender los efectos del genotipo, densidad de plantas y el sistema de siembra sobre el rendimiento, a fin de canalizar mejor las estrategias del manejo agronómico tendientes a incrementar la productividad del cultivo.
En la región de los valles altos de México no se tiene registro de trabajos donde se comparen estos factores, por lo que resulta importante dilucidar su efecto sobre los componentes fisiológicos y numéricos del rendimiento que permitan optimizar las prácticas de manejo tendientes a incrementar los niveles de producción de este cultivo y ampliar su adopción en zonas agrícolas potenciales. Con base en lo anterior, se llevó a cabo el presente trabajo con el objetivo de evaluar el efecto de dos sistemas de siembra, tres densidades y dos genotipos sobre los componentes fisiológicos y numéricos del rendimiento de semilla en el cultivo de canola.
Materiales y métodos
Ubicación del trabajo experimental
El experimento se realizó en el ciclo invierno-primavera 2019-2020 en la Facultad de Ciencias Agrícolas, dependiente de la Universidad Autónoma del Estado de México, localizada a 18 km al norte de la ciudad de Toluca, con coordenadas geográficas de 19° 15’ 33’’ de latitud norte, 99° 39’ 38’’ de longitud oeste y a una altitud de 2 640 msnm. El clima predominante es del tipo C (w2) (w) b (i) que corresponde a un clima templado subhúmedo, el más húmedo de los subhúmedos, con lluvias en verano y escasa precipitación pluvial en invierno (5%), poca oscilación térmica, temperatura media anual de 12.8 ºC y precipitación promedio anual de 900 mm.
El suelo predominante de la región es de tipo vertisol pélico de origen volcánico, presenta un horizonte mineral con un bajo contenido de materia orgánica que va del 1.01 a 2.36%. El color del horizonte superficial en seco es café grisáceo obscuro o gris obscuro con contenidos de arcilla de 20 a 36.4%. En el perfil del suelo se puede distinguir un horizonte con disturbios de labranza presentando compactaciones por piso de arado, la cantidad de materia orgánica es muy baja que va desde los 0.07 a 1.01 (Gil et al., 2014).
Tratamientos y diseño experimental
Se evaluaron dos sistemas de siembra SCDH: sistema cama doble hilera (24 parcelas) y SCP: sistema cama plana (24 parcelas). Los tratamientos derivados del arreglo factorial de dos genotipos de canola Hyola 61 (híbrido), Bioaureo 2486 (polinización abierta) y tres densidades, se establecieron bajo un diseño de bloques completos al azar con cuatro repeticiones en cada uno de los sistemas (dos experimentos en serie) considerando los dos sistemas de siembra.
Condiciones generales del experimento
La siembra se realizó el día 07 de diciembre de 2019 en dos sistemas de siembra, el primero conformado por parcelas de 3 m de longitud con tres camas separadas a 80 cm con dos hileras cada una separada a 20 cm (7.2 m2) sistema cama doble hilera (SCDH), mientras que el segundo contó con parcelas de 5 hileras de 3 m de longitud y separadas entre sí a 0.3 m (4.8 m2) sistema cama plana (SCP). Se utilizaron tres densidades de siembra 50, 75 y 90 semillas m-2. Para garantizar las tres densidades de siembra, la semilla se colocó en cintas de papel biodegradable a una distancia que garantizara la densidad requerida. El cultivo se mantuvo con humedad a capacidad de campo durante su ciclo y sin restricciones nutricionales por lo que se fertilizó con fórmula de 150-60-30 (NPK), utilizando como fuentes de nitrógeno a la urea, fósforo al superfosfato de calcio triple y al cloruro de potasio como fuente de este elemento.
Producción de biomasa, rendimiento y sus componentes
En madurez fisiológica se cortaron las plantas en un metro lineal del surco central de cada parcela y éstas fueron separadas en ramas y tallo principal, separando las silicuas de cada estrato. La materia seca de cada estrato (ramas, tallo principal y silicuas) se determinó después de secar en una estufa de aire forzado durante 72 h a 60 °C. Las silicuas de cada estrato se contaron y se trillaron por separado para obtener el peso seco de semilla (rendimiento). Se determinó el número de silicuas, el número de semillas por silicua y el número de semillas por m2. El peso de 1 000 semillas se determinó al contar y pesar 1 000 semillas de cada unidad experimental.
Análisis estadístico
Las variables de respuesta del experimento fueron sometidas a un análisis de varianza combinado y cuando la prueba de F de los análisis de varianza resultó significativa, se procedió a realizar la prueba de medias utilizando mediante la diferencia mínima significativa honesta (DMSH) o prueba de Tukey a un nivel de significancia del 5% (Palaniswamy y Palaniswamy, 2006), utilizando el paquete estadístico R para Windows versión 4.0.5 en base al siguiente modelo:
.
En donde:
es la media general; = es el efecto del i-ésimo sistema;
es el efecto del j-ésimo bloque dentro de sistemas y fue el termino de error usado para probar sistemas;
es el efecto de la k-ésima densidad;
es el efecto del l-ésimo genotipo;
es efecto de la interacción de la k-ésima densidad en el i-ésimo sistema;
es el efecto del i-ésimo sistema en el l-ésimo genotipo;
es el efecto de la k-ésima densidad en el l-ésimo genotipo;
es la interacción del i-ésimo sistema en la k-ésima densidad en el l-ésimo genotipo y
es el termino de error general. Las relaciones entre variables fueron determinadas mediante regresión lineal y la asociación entre las mismas mediante análisis de correlación (Jandel, 1991).
Resultados y discusión
Condiciones climáticas
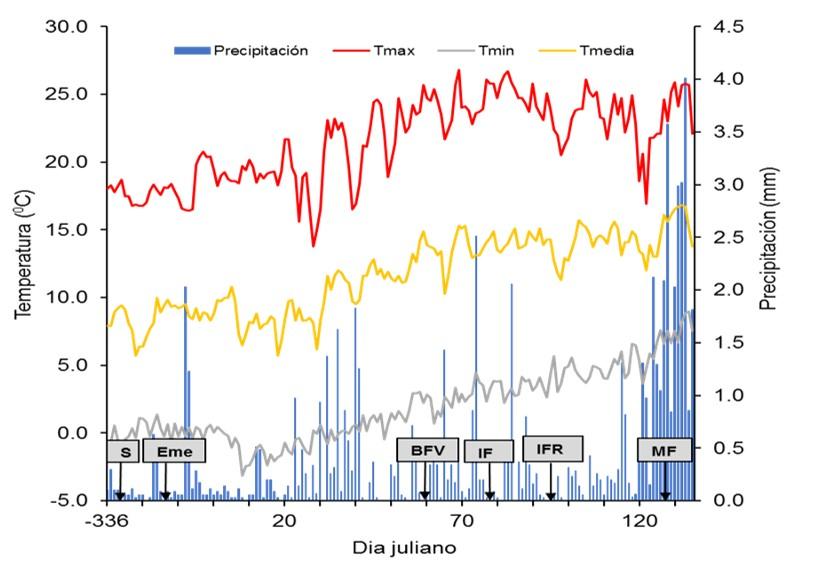
Las condiciones climáticas durante el desarrollo de los experimentos se presentan en la Figura 1, en la cual se observa que la emergencia ocurrió a los 14 días posteriores a la siembra. Por otro lado, se puede apreciar que el inicio del periodo reproductivo en ambos genotipos coincidió con temperaturas favorables para el cultivo. Cabe señalar, que la precipitación durante el periodo reproductivo fue muy escasa, pero esto no fue una limitante dado que el cultivo fue establecido bajo condiciones de riego. Hacia la madurez fisiológica se observó un ascenso en los niveles de precipitación registrados, lo cual no repercutió sobre el desarrollo del experimento. En general los factores como precipitación y temperatura fueron favorables para el desarrollo del experimento.
Los resultados del análisis de varianza que se presentan en el Cuadro 1, muestran que para el factor sistema de siembra únicamente se observaron diferencias altamente significativas (p< 0.01) para las variables biomasa, peso de 1 000 semillas y número de ramas. Por otra parte, para genotipos se observaron diferencias altamente significativas (p< 0.01) en biomasa, peso de 1000 semillas y número de ramas y únicamente se observaron diferencias significativas (p< 0.05) para índice de cosecha. Para las interacciones no se observaron efectos significativos para la mayoría de las variables con excepción de la interacción sistema x densidad x genotipo la cual mostró efectos altamente significativos (p< 0.01) para número de silicuas. Finalmente, los coeficientes de variación oscilaron entre 1.2 y 20.8 que corresponden a las variables peso de 1 000 semillas y biomasa a madurez fisiológica dichos coeficientes se consideran aceptables en función de la naturaleza del experimento.
Figura 1. Condiciones agroclimáticas ocurridas durante la conducción del trabajo experimental. (S= siembra; Eme= emergencia; BFV= botón floral visible; IF= inicio de la floración; IFR= inicio de la fructificación; MF= madurez fisiológica).
Cuadro 1. Valores de F y su significancia estadística para las variables evaluadas en los dos sistemas de siembra, tres densidades y dos genotipos de canola en Toluca, México.
Fuente de variación | gl | REND (g m-2) | BIOM (g m-2) | IC | NG (m2) | P1000S (g) | NS (m2) | NSPS | NR |
Sistemas (S) | 1 | 0.4 ns | 13.2 ** | 3 ns | 0.1 ns | 24.7 ** | 3.6 ns | 1.3 ns | 15.7 ** |
Bloques (sistema) | 6 | 2.5 * | 0.9 ns | 0.6 ns | 2.5 * | 0.1 ns | 0.9 ns | 1.4 ns | 2.5 * |
Densidades (D) | 2 | 9.7 ** | 7.6 ** | 1.1 ns | 9.3 ** | 0 ns | 108.1 ** | 0.4 ns | 22 ** |
Genotipos (G) | 1 | 1.3 ns | 8.2 ** | 4.6 * | 0.9 ns | 7.4 ** | 2.5 ns | 0 ns | 20.8 ** |
S x D | 2 | 0.2 ns | 0.6 ns | 0.9 ns | 0.1 ns | 0.6 ns | 2.9 ns | 1.5 ns | 3 ns |
S x G | 1 | 3.5 ns | 0.4 ns | 0.5 ns | 3.5 ns | 0 ns | 0.3 ns | 0 ns | 1.3 ns |
D x G | 2 | 0.3 ns | 0.9 ns | 1.1 ns | 0.3 ns | 0.2 ns | 0.7 ns | 0.5 ns | 2.5 ns |
S x D x G | 2 | 0.1 ns | 0.5 ns | 0.9 ns | 0.1 ns | 1.2 ns | 13.6 ** | 0.3 ns | 1 ns |
CV (%) | 17.4 | 20.8 | 20 | 17.5 | 1.2 | 8.6 | 14.1 | 10 |
REND= Rendimiento; BIOM= biomasa a madurez; IC= índice de cosecha; NG= número de semillas por m2; P1 000S= peso de 1 000 semillas; NS= número de silicuas por m2; NSPS= número de semillas por silicua; NR= número de ramas; *= significativo al 0.05; **= significativo al 0.01; ns= no significativo y gl= grados de libertad.
En el Cuadro 2, se aprecia la comparación de medias para las variables evaluadas, donde el sistema cama plana (SCP) fue significativamente superior al sistema cama doble hilera (SCDH) en producción de biomasa, peso de 1 000 semillas y número de ramas por planta, para el resto de los caracteres ambos sistemas de siembra resultaron estadísticamente iguales. La superioridad del sistema cama plana sobre el sistema de cama doble hilera radica en que en el primero la separación de las hileras fue de 30 cm, mientras que en el segundo la separación fue de 20 cm, pero en camas separadas a 80 cm. Evidencias en la literatura muestran que un menor espaciamiento entre hileras produce un mayor rendimiento de semilla como consecuencia de mayor número de silicuas y semillas por m2 (Shahin y Valiollah, 2009; Uzun et al., 2012).
Cuadro 2. Valores medios de las variables evaluadas en dos sistemas de siembra, tres densidades y dos genotipos de canola en Toluca, México.
REND (g m-2) | BIOM (g m-2) | IC | NG (m2) | P1000S (g) | NS (m2) | NSPS | NR | |
Sistema | ||||||||
SCP | 493.3 a | 2 609.7 a | 0.19 a | 128549 a | 3.8 a | 2 117.8 a | 19.6 a | 5.5 a |
SCDH | 467.6 a | 2 115.6 b | 0.2 a | 124135 a | 3.7 b | 2 216.7 a | 20.7 a | 4.5 b |
Media | 480.4 | 2 362.6 | 0.15 | 126342 | 3.7 | 2 167.2 | 20.1 | 50 |
Densidad | ||||||||
D90 | 552.4 a | 2 692.7 a | 0.2 a | 145007 a | 3.8 a | 2 541.8 a | 19.7 a | 4.4 c |
D75 | 463.8 b | 2 381b a | 0.18 a | 122060 b | 3.8 a | 2 343.1 b | 20.1 a | 5 b |
D50 | 425.2 b | 2 014.2 b | 0.2 a | 111958 b | 3.8 a | 1 616.9 c | 20.6 a | 5.6 a |
Media | 480.4 | 2 362.6 | 0.16 | 126341 | 3.8 | 2 167.2 | 20.1 | 5 |
Genotipo | ||||||||
Hyola 61 | 466.3 a | 2 158.6 b | 0.21 a | 123181 a | 3.7 b | 2 124.3 a | 20.2 a | 4.7 b |
Bioaureo 2486 | 494.6 a | 2 566.6 a | 0.18 b | 129502 a | 3.8 a | 2 210.2 a | 20 a | 5.3 a |
Media | 480.4 | 2 362.6 | 0.19 | 126341 | 3.7 | 2 167.2 | 20.1 | 5 |
REND= rendimiento; BIOM= biomasa a madurez; IC= índice de cosecha= NG= número de semillas por m2; P1 000S= peso de 1 000 semillas; NS= número de silicuas por m2; NSPS= número de semillas por silicua; NR= número de ramas. Medias con la misma letra dentro de columnas no difieren significativamente entre si, a un nivel de significancia del 0.05 de la prueba de Tukey.
Incrementos en la densidad de plantas se ven reflejados en una mayor producción de biomasa y rendimiento de semillas, como consecuencia de un mayor número de semillas por m2 (Shahin and Valiollah, 2009; Uzun et al., 2012). Los datos revelan que la D90 (90 plantas m-2) fue estadísticamente superior a las D75 (75 plantas m-2) y D50 (50 plantas m-2) en la mayoría de las variables evaluadas con excepción del índice de cosecha, peso de 1 000 semillas y número de semillas por silicua.
El efecto genotípico sobre la densidad de población en canola ha sido analizado en muchos estudios, en donde las diferencias en rendimiento de semilla han sido mínimas (Różyło y Pałys, 2014), mientras que otros mencionan que las diferencias en rendimiento pueden estar dadas por la diferente estructura de la planta (Waseem et al., 2014). En este sentido nuestros datos muestran que Hyola 61 fue estadísticamente superior (14%), a Bioaureo 2486 en índice de cosecha, mientras que Bioaureo 2486 fue estadísticamente superior en producción biomasa (18%), peso de 1 000 semillas (3%) y número de ramas (11%) a Hyola 61 (Cuadro 2). Las diferencias en algunas características entre ambos cultivares pueden estar dadas por la diferente estructura de planta que existe entre ellas ya que Hyola 61 produce menos ramas que Bioaureo 2486.
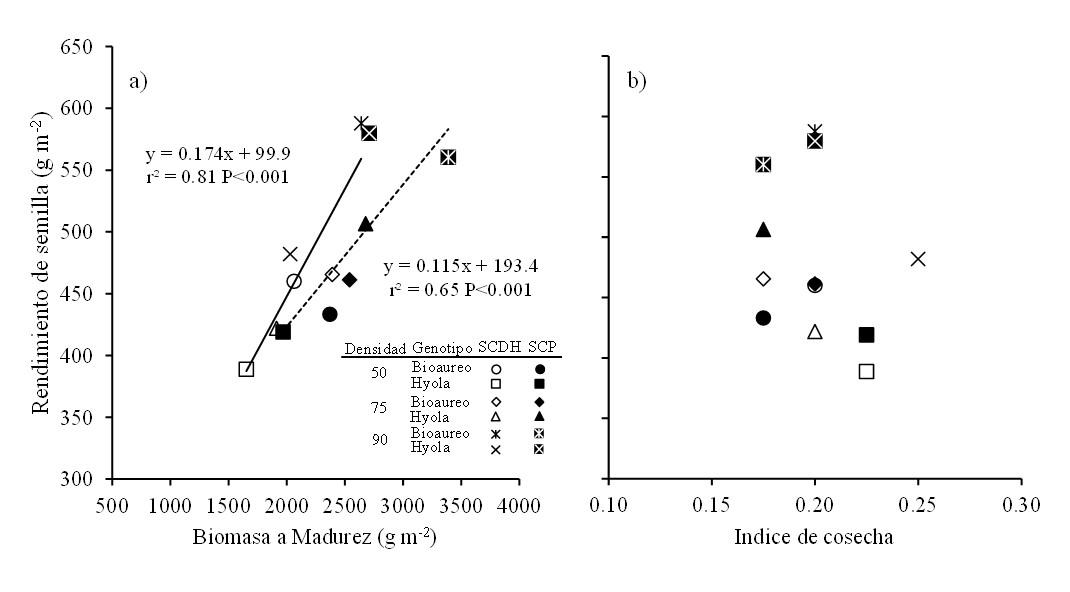
La producción de biomasa a madurez y el índice de cosecha, son características muy importantes para lograr altos rendimientos en los cultivos de grano y particularmente en canola (Zhang y Flottmann, 2016). La contribución de ambos caracteres al rendimiento de semilla depende fuertemente del tipo de cultivar (hibrido o polinización abierta), pero por lo general la producción de biomasa explica en gran medida los cambios en el rendimiento de semilla (Li et al., 2016). Los datos indican que, al considerar los sistemas de siembra, densidades y genotipos, el rendimiento de semilla fue explicado principalmente por los cambios observados en la producción de biomasa a madurez (r2= 0.81, p< 0.001) y no por el índice de cosecha (Figura 2), este último al parecer fue más estable, como ha sido reportado en otros cultivos de grano como triticale (Estrada et al., 2012) y trigo (Parry y Hawkesford, 2010).
Se ha establecido que el distanciamiento entre hileras es un factor de manejo muy importante ya que afecta el rendimiento de semilla y los componentes del rendimiento en planta individual en canola (Uzun et al., 2012; Różyło y Pałys, 2014). Nuestros datos revelaron que cuando se consideran densidades y genotipos el sistema cama plana (cinco hileras a 30 cm), obtuvo una mayor producción de biomasa respecto al sistema cama doble hilera (Cuadro 3; Figura 2a).
Figura 2. Relaciones entre el rendimiento de semilla y la biomasa producida en madurez (a) y con el índice de cosecha (b) para dos cultivares de canola cultivados bajo tres densidades de plantas y en dos sistemas de siembra en Toluca, México. Línea continua y discontinua en el gráfico son los modelos de regresión ajustados para SCDH y SCP, respectivamente.
Por otro lado, incrementos en la densidad de siembra aumentaron significativamente el rendimiento de semilla en ambos sistemas de siembra, siendo incrementos de 22 y 20% al pasar de 50 a 90 plantas m-2 en Bioaureo 2486 y Hyola 61, respectivamente para el sistema cama doble hilera. En tanto que, en el sistema cama plana los incrementos en rendimiento al pasar de 50 a 90 plantas m-2 fue de 23 y 28% para Bioaureo 2486 y Hyola 61, respectivamente (Cuadro 3).
Cuadro 3. Valores medios de las variables evaluadas para sistema, densidad y genotipo en el cultivo de canola en Toluca, México.
Densidad | Genotipo | REND (g m-2) | BIOM (g m-2) | IC | NG (m2) | |||||||
SCDH | SCP | SCDP | SCP | SCDP | SCP | SCDP | SCP | |||||
50 | Bioaureo | 460bA | 433bA | 2 063aA | 2370bA | 0.2aA | 0.17aB | 121992aA | 112509bA | |||
Hyola | 388bA | 419bA | 1 653bA | 1969cA | 0.22aA | 0.22aA | 103026bA | 110304bA | ||||
75 | Bioaureo | 465aA | 461aA | 2 392aA | 2540bA | 0.17bB | 0.2aA | 122524aA | 119953bA | |||
Hyola | 422bA | 506aA | 1 913aB | 2677aA | 0.2aA | 017aB | 113408bA | 132354aA | ||||
90 | Bioaureo | 587aA | 560aA | 2 641aB | 3390aA | 0.2aA | 0.17aB | 155491aA | 144541aA | |||
Hyola | 482aA | 579aA | 2 029aB | 2709aA | 0.25aA | 0.2aB | 128366aA | 151628aA | ||||
467A | 493A | 2 115B | 2609A | 0.2A | 0.19A | 124135A | 128549A | |||||
P1000S(g) | NS (m2) | NSPS | NR | |||||||||
50 | Bioaureo | 3.8aA | 3.9aA | 1 724A | 1 538cB | 21.5aA | 19.2aA | 5.7aA | 6aA | |||
Hyola | 3.8aA | 3.8aA | 1 662A | 1 543cA | 22.1aA | 19.8aA | 5aB | 5.7aA | ||||
75 | Bioaureo | 3.8aA | 3.9aA | 2 620B | 2 119bA | 22.3aA | 19.0aB | 4.5bB | 6aA | |||
Hyola | 3.7aA | 3.8aA | 2 292A | 2 340bA | 20aA | 19.1aA | 4.2bB | 5.5aA | ||||
90 | Bioaureo | 3.8aA | 3.9aA | 2 385B | 2 873aA | 18.6aA | 19.9aA | 4.7aA | 5.2bA | |||
Hyola | 3.8aA | 3.8aA | 2 616B | 2 291bA | 20aA | 20.4aA | 3.2cA | 4.5bB | ||||
3.7A | 3.8A | 2 216A | 2 117A | 20.7A | 19.A | 4.5B | 5.5A | |||||
REND= Rendimiento; BIOM= biomasa a madurez; IC= índice de cosecha; NG= número de semillas por m2; P1 000S= peso de 1 000 semillas; NS= número de silicuas por m2; NSPS= número de semillas por silicua; NR= número de ramas; S= sistema; D= densidad; G= genotipo. Medias con la misma letra mayúscula no son diferentes entre sistemas y misma letra minúscula dentro de columnas las medias no difieren significativamente entre si, a un nivel de significancia del 0.05 de la prueba de Tukey.
Efectos positivos en la reducción de la distancia entre hileras han sido reportados en numerosos estudios, por ejemplo, Shahin y Valiollah (2009), al evaluar tres distanciamientos entre hileras (12, 18 y 24 cm) encontraron incrementos significativos en el rendimiento de semilla en hileras separadas a 12 cm. Por otro lado, Uzun et al. (2012) al evaluar distintos espaciamientos entre plantas (5, 10, 15 y 20 cm) y entre surcos (10, 20, 30 y 40 cm) encontraron un mayor rendimiento con distanciamientos entre plantas de 5 a 10 cm y separación entre surcos de 10 cm. En la misma forma, Różyło y Pałys (2014) al evaluar distintos espaciamientos entre surcos encontraron que una mayor separación disminuyó tanto la producción de biomasa como el rendimiento por m2. Recientemente French et al. (2016), al evaluar densidades entre 7 y 180 plantas m-2, encontraron que la densidad promedio óptima fue de 32 plantas m-2 y conforme se incrementaba la densidad existía un efecto positivo sobre el rendimiento.
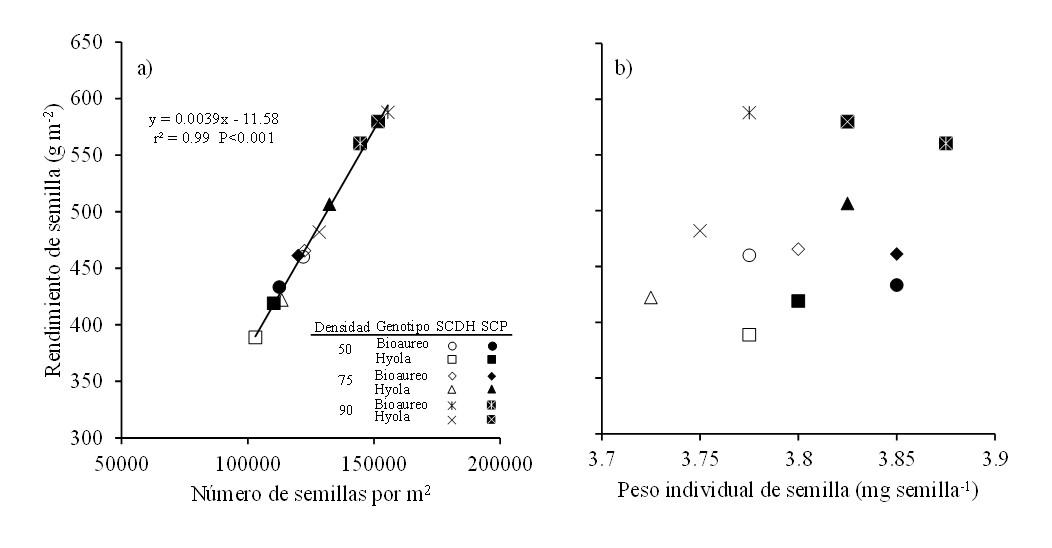
El análisis de los componentes numéricos del rendimiento en los cultivos de grano ha mostrado que el componente más importante en la determinación del rendimiento es el número de granos por m2 (Cheng-dong et al., 2019). Como era de esperarse, los cambios en el rendimiento de semilla al considerar sistemas de siembra, densidades y cultivares fueron explicados por el número de semillas por m2 (r2= 0.99, p< 0.001; Figura 3a), dado que para el peso individual de semilla no se observó ningún grado de asociación (Figura 3b). En ambos cultivares se observó que el incremento en la densidad de siembra afectó positivamente el número de semillas por m2 en el sistema cama doble hilera, sin embargo, esto no afectó estadísticamente el peso individual de semilla (Figura 3b).
Figura 3. Relaciones entre el rendimiento de semilla y el número de semillas por m2 (a) y con el peso individual de semilla (b) para dos cultivares de canola cultivados bajo tres densidades de población y en dos sistemas de siembra en Toluca, México. Línea continua en el gráfico representa el modelo de regresión ajustado para ambos sistemas.
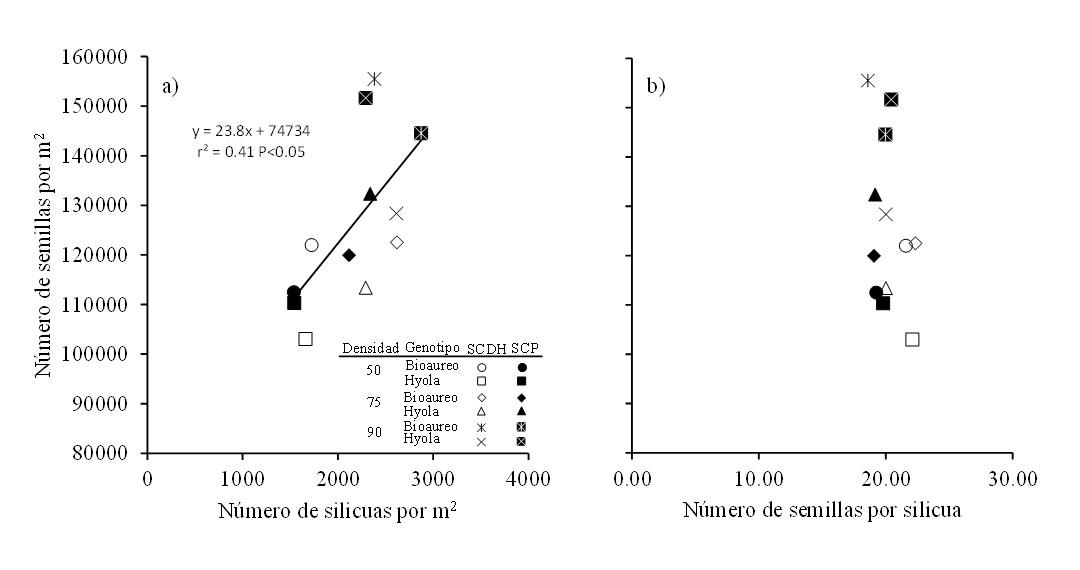
Cabe resaltar que el número de semillas por m2 tuvo una asociación moderada y positiva con el número de silicuas por m2 (r2= 0.41 p< 0.05) (Figura 4a) cuando se consideró el efecto conjunto de sistemas de siembra, densidades y cultivares, mientras que para un mismo número de semillas por silicua se observaron cambios en el número de semillas por m2 (Figura 4b). Estos resultados resaltan la importancia del número de silicuas como una característica asociada al rendimiento de semilla a través del número de semillas (Shahin y Valiollah 2009). El número se silicuas por planta depende del número de ramas, número de racimos florales y de cuantas de estas silicuas cuajen (Shahin y Valiollah, 2009; Assefa et al., 2018) y es un carácter que compensa pérdidas en rendimiento por baja densidad en ambientes potencialmente buenos, debido a un mayor número de silicuas por rama productiva cuando la densidad de siembra es reducida (Assefa et al. 2018).
Figura 4. Relaciones entre el número de semillas por m2 con el número de silicuas por m2 (a) y con el número de semillas por silicua (b) para dos cultivares de canola cultivados bajo tres densidades de población y en dos sistemas de siembra en Toluca, México. Línea continua en el gráfico representa el modelo de regresión ajustado para ambos sistemas.
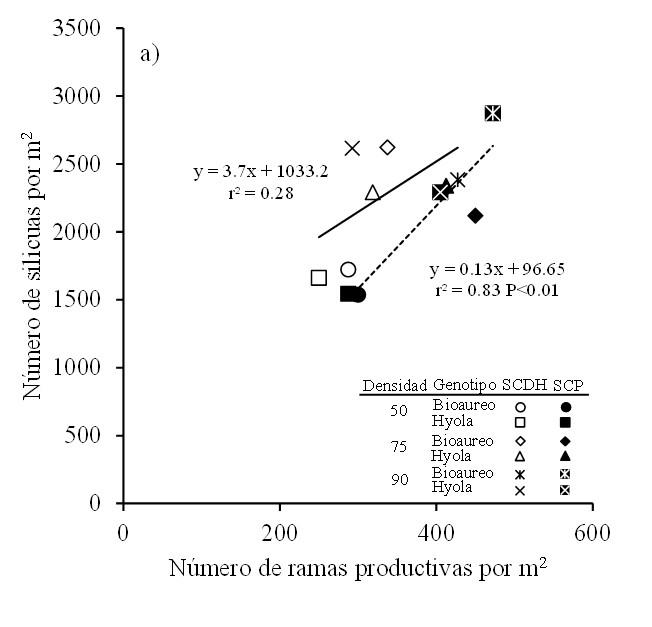
El número de ramas productivas en canola es una característica que está asociada grandemente con el rendimiento de semilla y también responde en forma significativa a los cambios en el ambiente y prácticas de manejo (Shahin y Valiollah, 2009; Hua et al., 2014). En este sentido, se observan reducciones en el número de ramas cuando se incrementa la densidad de siembra (Uzun et al., 2012; Vincze, 2017) o cuando existen retrasos en la fecha de siembra (Hua et al., 2014; Vincze, 2017). Nuestros datos revelan que el número de silicuas por m2 estuvo asociado con los cambios observados en el número de ramas productivas, principalmente en el sistema cama plana (r2= 0.83, p< 0.01) (Figura 5).
Figura 5. Relación entre el número de silicuas por m2 y el número de ramas por m2 para dos cultivares de canola cultivados bajo tres densidades de población y en dos sistemas de siembra en Toluca, México. Línea continua y discontinua en el gráfico son los modelos de regresión ajustados para SCDH y SCP, respectivamente.
El rendimiento se semilla se asoció positivamente con la producción de biomasa en madurez fisiológica, con el número de semillas por m2 y con el número de silicuas por m2 (Cuadro 4) lo cual concuerda con numerosos estudios en la literatura que indican que estos atributos son definitorios del rendimiento de semilla en canola (Shahin y Valiollah, 2009; Uzun et al., 2012; Mamun et al., 2014; Yang et al., 2014). Por otro lado, la biomasa en madurez fisiológica se correlacionó de forma positiva con el número de semillas, con el peso de 1 000 semillas y con el número de silicuas por m2, mientras que una asociación negativa fue observada con el índice de cosecha.
Como se esperaba el número de semillas por m2 se correlacionó positivamente con el número de silicuas por m2, lo cual indica la gran influencia del número de silicuas en la determinación del número de semillas y el rendimiento en canola, coincidiendo con evidencias previas en la literatura (Shahin y Valiollah, 2009; Kazemeini et al., 2010). Por otra parte, existió una asociación negativa entre el número de silicuas por m2 y el número de ramas. Esto es un aspecto muy importante para considerar cuando se varía la densidad, ya que el número de ramas productivas tiende a afectarse tanto por densidad como por retrasos en la fecha de siembra (Kazemeini et al., 2010; Vincze, 2017; Assefa et al., 2018). El peso de 1 000 semillas presentó una asociación positiva con el número de ramas.
Cuadro 4. Coeficientes de correlación entre los caracteres evaluados.
REND | BIOM | IC | NG | P1000S | NS | NGPS | |
Biom | 0.708** | ||||||
IC | 0.029 | -0.453** | |||||
NG | 0.997** | 0.68** | 0.048 | ||||
P1000G | 0.188 | 0.473** | -0.257 | 0.117 | |||
NS | 0.468** | 0.502** | -0.098 | 0.468** | 0.025 | ||
NGPS | -0.106 | -0.178 | -0.067 | -0.091 | -0.211 | -0.02 | |
NR | -0.099 | 0.14 | -0.218 | -0.13 | 0.448** | -0.444** | -0.06 |
**= altamente significativo al 0.01. REND= rendimiento de semilla (g m-2); BIOM= biomasa en madurez fisiológica (g m-2); IC= índice de cosecha; NG= número de semillas por m2; P1 000 semillas= peso de 1 000 semillas (g); NS= número de silicuas por m2; NGPS= número de semillas por silicua; NR= número de ramas productivas por planta.
Conclusiones
En términos de componentes fisiológicos, el rendimiento de semilla fue explicado por los cambios observados en la producción de biomasa a madurez y no por el índice de cosecha. El mayor rendimiento de semilla se obtuvo con el sistema cama plana, el genotipo Bioaureo 2486 y una densidad de 90 plantas m-2. Con el sistema cama plana (cinco hileras a 30 cm), se obtuvo una mayor producción de biomasa respecto al sistema cama doble hilera. Los dos cultivares mostraron diferencias en su estructura de planta, Hyola 61 produjo menos ramas que Bioaureo 2486.
Agradecimientos
El autor G. García-Hernández desea agradecer al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca que le permitió terminar sus estudios de Maestría en Ciencias Agropecuarias y Recursos Naturales. El presente trabajo fue parte del proyecto ‘Atributos ecofisiológicos del rendimiento y producción de biomasa en cultivos de grano modificados por la densidad de población y sistema de siembra’ con clave de registro en la UAEMex.
Literatura citada
Assefa, Y.; Carter, P.; Hinds, M.; Bhalla, G.; Schon, R.; Jeschke, M.; Paszkiewicz, S.; Smith, S. and Ciampitti, I. A. 2018. Analysis of long-term study indicates both agronomic optimal plant density and increase maize yield per plant contributed to yield gain. Sci. Rep. 8(1):1-11. https://doi.org/10.1038/s41598‐018‐23362‐x.
Cheng-dong, H.; Quan-quing. L.; Xiao-lin, L. and Chao-chun, Z. 2019. Effect of intercropping on maize grain yield and yield components. J. Integrative Agric. 18(8):1690-1700. Doi:10.1016/s2095-3119(19)62648-1.
CONASIPRO. 2019. Bases de datos oleaginosas mundial y nacional 1980-2018. http://www.oleaginosas.org/cat-69.shtml#51.
Estrada, C. G.; Slafer, G. A. and Miralles, D. J. 2012. Differences in yield, biomass and their components between triticale and wheat grown under contrasting water and nitrogen environments. Field Crops Res. 128:167-179. https://doi.org/10.1016/j.fcr.2012.01.003.
FAOSTAT. 2018. Organización de las Naciones Unidas para la Agricultura y la Alimentación. Base de datos estadísticos. http://www.fao.org.
French, R. J.; Seymour, M. and Malik, R. S. 2016. Plant density response and optimum crop densities for canola (Brassica napus L.) in Western Australia. Crop Pasture Sci. 67(4):397-408. https://doi.org/10.1071/CP15373.
Gan, Y.T.; Harker, K. N.; Kutcher, H. R.; Gulden, R. H.; Irvine, B.; May, W. E. and O’Donovan, J. T. 2016. Canola seed yield and phenological responses to plant density. Can. J. Plant Sci. 96(1):151-159. https://doi.org/10.1139/cjps-2015-0093.
Gil, H.; Martínez, C. G y Estrada, G. 2014. Impacto del sistema de labranza y dosis de nitrógeno en el rendimiento y calidad nutricional de forraje de avena. Rev. Mex. Cienc. Agríc. 6(5):951-964.
Hosseini, M. N.; Alizadeh, H. M. and Ahmadi, H. M. 2006. Effects of plant density and nitrogen rates on the competitive ability of canola (Brassica napus L.) against weeds. J. Agric. Sci. Technol. 8(4):281-291.
Hua, S.; Lin, B.; Hussain, N.; Zhang, Y.; Yu, H.; Ren, Y.; Ding, H. and Zang, D. 2014. Delayed planting affects seed yield, biomass production, and carbohydrate allocation in canola (Brassica napus). Int. J. Agric. Biol. 16(4):671-680.
Jandel, 1991. Table Curve v. 3.0. User’s Manual Versión 3.0. AISN Software. Jandel Scientific, Corte Madera, CA.
Kazemeini, A.; Edalat, M.; Shekoofa, A. and Hamidi, R. 2010. Effects of nitrogen and plant density on rapeseed (Brassica napus L.) yield and yield components in Southern Iran. Rev. Cienc. Aplicadas. 10(14):1461-1465. Doi:10.3923/jas.2010.1461.1465.
Kirkegaard, A. J.; Lilley, M. J. and Morrison, M. J. 2016. Drivers of trends in Australian canola productivity and prospects. Crop Pasture Sci. 67(4):i-ix. https://doi.org/10.1071/CPv67n4-FO.
Li, S. Y.; Yu, C. B.; Zhu, S.; Xie, L. H.; Hu, X. J.; Liao, X.; Liao, S. X. and Che, Z. 2014. High planting density benefits to mechanized harvest and nitrogen application rates of oilseed rape (Brassica napus L.). Soil Sci. Plant Nutr. 60(3):384-392. Doi:10.1080/00380768. 2014.895417.
Li, X.; Li, Q.; Yang, T.; Nie, Z.; Chen, G. and Hu, L. 2016. Responses of plant development, biomass and seed production of direct sown oilseed rape (Brassica napus L.) to nitrogen application at different stages in Yangtze River Basin. Field Crops Res. 194:12-20. Doi: 10.1016/j.fcr.2016.04.024.
Mamun, F.; Ali, M. H.; Chowdhury, I. F.; Hasanuzzaman, M. and Matin, M. A. 2014. Performance of rapeseed and mustard varieties grown under different plant density. Sci. Agric. 4(2):70-75. Doi: 10.15192/PSCP.SA.2014.4.2.7075.
Mobasser, H. R.; Shojaee-Ghadikolaee, M.; Nasiri, M.; Daneshian, J.; BarariTari, D. and Pourkalhor, H. 2008. Effect of nitrogen rates and plant density on the agronomic traits of canola (Brassica napus L.) in paddy field. Asian J. Plant Sci. 7(2):233-236. Doi:10.3923/ajps.2008.233.236.
Palaniswamy, U. R. and Palaniswamy, K. M. 2006. Handbook of statistics for teaching and research in plant and crop science. The Harworth Press, Inc., New York. 624 p.
Parry, M. A. J. and Hawkesford, M. J. 2010. Food security: increasing yield and improving resource use efficiency. Proceed. Nutr. Soc. 69(04):592-600. https://doi.org/10.1017/ S0029665110003836.
Rathke, G. W.; Behrens, T. and Diepenbrock, W. 2006. Integrated management strategies to improve seed yield; oil content and nitrogen efficiency of winter oilseed rape (Brassica napus L.): a review. Agric. Ecosys, Environ. 117(2-3):80-108. https://doi.org/10.1016/ j.agee.2006.04.006.
Rondanini, D. R.; Menéndez, Y. C.; Gómez, N. V.; Miralles, D. J. and Botto, J. F. 2017. Vegetative plasticity and floral branching compensate low plant density in modern spring rapeseed. Field Crop Res. 210:104-113. https://doi.org/10.1016/j.fcr.2017.05.021.
Różyło, K. and Pałys, E. 2014. New oilseed rape (Brassica napus L.) varieties-canopy development, yield components, and plant density. Section B - Soil & Plant Sci. Acta Agric. Scandinavica 64(3):260-266.
Shahin, Y. and Valiollah, R. 2009. Effects of row spacing and seeding rates on someagronomical traits of spring canola (Brassica napus L.) cultivars. J. Cent. Eur. Agric. 10(1):115-212.
Uzun, B.; Yol, E. and Furat, S. 2012. The influence of row and intra-row spacing to seed yield and its components of winter sowing canola in the true Mediterranean type of environment. Bulg. J. Agric. Sci. 18(1):83-91.
Vincze, E. 2017. The effect of sowing date and plant density on yield elements of different winter oil seed rape (Brassica napus var. napus F. biennis L.) genotypes. Columella. 1(4):21-25. Doi: 10.18380/szie.colum.2017.4.1.suppl.
Wang, R.; Cheng, T. and Hu, L. 2015. Effect of wide-narrow row arrangement and plant density on yield and radiation use efficiency of mechanized direct-seeded canola in Central China. Field Crops Res. 172:42-52. Doi:10.1016/j.fcr. 2014.12.005.
Waseem, M.; Baloch, D. and Khan, I. 2014. Influence of various row spacing on the yield and yield components of Raya Anmol and Faisal Canola under coastal climatic conditions of Lasbela. Am. J. Plant Sci. 5(15):2230-2237. Doi:10.4236/ajps.2014.515237.
Yang, C.; Gan, Y.; Harker, K. N.; Kutcher, H. R.; Gulden, R.; Irvine, B. and May, W. E. 2014. Up to 32% yield increase with optimized spatial patterns of canola plant establishment in western Canada. Agron. Sustain. Dev. 34:793-801. https://doi.org/10.1007/s13593-014-0218-5.
Zhang, H. and Flottmann, S. 2016. Seed yield of canola (Brassica napus L.) is determined primarily by biomass in a high-yielding environment. Crop Pasture Scie. 67(4):369-380. https://doi.org/10.1071/CP15236.