Revista Mexicana de Ciencias Agrícolas special publication number 23 September 28 - November 11, 2019
DOI: https://doi.org/10.29312/remexca.v0i23.2023
Article
Characterization of cultivated varieties of Capsicum pubescens
in Michoacán, Mexico
Ana Karen Escalera-Ordaz1
Héctor Guillén-Andrade2§
Ma. Blanca Nieves Lara-Chávez2
Clemente Lemus-Flores1
Javier German Rodríguez-Carpena1
Roberto Valdivia-Bernal1
1Academic Unit of Agriculture-Autonomous University of Nayarit. Tepic-Compostela Highway km 9, Xalisco, Nayarit, Mexico. CP. 63780. Tel. 5212112478. (ana.karen.escalera.ordaz@gmail.com; clemus23@gmail.com; germencillo@yahoo.com.mx; beto49-2000@yahoo.com.mx). 2Faculty of Agrobiology President Juárez-Advanced Research Unit in Agrobiotechnology-Michoacana University of San Nicolás de Hidalgo. Paseo Gral. Lázaro Cárdenas and Berlin s/n, Col. Viveros, Uruapan, Michoacán, Mexico. CP. 60170. Tel. 452 5236474. (chavez12001@yahoo.com.mx).
§Corresponding author: hguillenandrade@gmail.com.
Abstract
The state of Michoacán has diverse homogeneous agro-ecological zones that favor the wide diversity of plant species, which includes the cultivation of chili, considered one of the main crops worldwide. The cultivation of C. pubescens has gained greater importance in recent years due to the increase in its production. This species has broad genetic variability that can be used in breeding programs. However, the variability in the state of Michoacán has not been sufficiently documented, so the genetic potential it represents is unknown. In the present work, the collection and morphological characterization of cultivated varieties of C. pubescens in different agroecological zones was proposed in order to evaluate the morphological variability and relate it to the different existing agro-ecological conditions. The in situ morphological characterization was made based on the descriptors proposed by IPGRI, AVRDC and CATIE and UPOV, the evaluation included 27 characteristics of leaf, fruit and seed. The results obtained indicate that there is a wide variability in the cultivated varieties of C. pubescens. Likewise, the fruit characteristics were identified as the most important to discriminate the varieties with potential to be used, as quality parameters, in breeding programs. Finally, no relationship was found between the morphological variability of C. pubescens and the sites of origin of the cultivated varieties.
Keywords: genetic resources, perón chili.
Reception date: June 2019
Acceptance date: August 2019
Introduction
The cultivation of chili is one of the most important horticultural crops in the world (de la Cruz-Lázaro et al., 2017). It is distinguished by the pungency of its fruits, mainly caused by three capsaicinoids: nordihydro-, dihydro- and capsaicin (Sánchez-Sánchez et al., 2010). It presents a wide variability of shapes, colors, aromas, flavors and sizes that constitute a valuable contribution of Mexico to the culture, food and gastronomy of the world since pre-hispanic times (Alonso et al., 2012; Hernández-Verdugo, 2014; Pérez et al., 2015), with great relevance in Mexican cuisine, being the main spice (Zegbe et al., 2012).
The chemical-food value is important because of the content of valuable antioxidant compounds in the prevention of chronic and degenerative diseases (López-López et al., 2015). Likewise, its economic value is increased in the process industry and extraction of dyes and oleoresins (Arreguin et al., 2015). In Mexico, the production of chili is 1 276 123 t ha-1, the main producing states are: Sinaloa, Sonora, Baja California Sur, Jalisco, San Luis Potosí, Veracruz, Tamaulipas, Guanajuato and Michoacán.
The latter contributes with a planted surface of 448 ha of the national surface and with a yield of 11 095 t ha-1 (SIAP, 2019). However, one of the main species of chili cultivated in the states of Michoacán, Veracruz, State of Mexico and Puebla is C. pubescens (Barrios-Puente et al., 2014), with great importance in national and foreign markets, and that its production has been increased by the high yields obtained, fruit quality and high profitability, the price fluctuates from $10.00 to $80.00 and is directly related to the season, size and fruit color.
Due to the above, there is a potential demand in growth for its consumption and production (Espinosa-Torres and Ramírez-Abarca, 2016). This species is characterized by being mainly rainfed, where the most significant months for harvesting are from August to December (Barrios-Puente et al., 2014). In Mexico, C. pubescens is commonly known as manzano chili, rocoto, canario or chili wax and in the state of Michoacán, it is recognized as perón chili (Espinosa-Torres and Ramírez-Abarca, 2016) and has wide genetic variability that can be used in genetic improvement programs as a result of the wide diversity of existing environments (Bobadilla-Larios et al., 2017).
In this state, there are diverse homogeneous agroecological zones, which are generally favorable for the development of avocado cultivation (Gutierréz-Contreras et al., 2010) and some favor the development of perón chili, since good adaptation is observed in areas of shade where warm air mixes with cold wind, under the avocado trees (SAGARPA, 2013).
However, C. pubescens is known only under cultivation and the lack of knowledge about diversity and phylogenetic relationships limit its genetic improvement (Pérez et al., 2015). In this sense, the related phytogenetic resources acquire great relevance because of the genetic potential they present and because they are the basis for obtaining improved varieties (Moreno-Pérez et al., 2011).
According to the above, there is a need to know the distribution, variability and genetics of the materials retained by the producers in their plots (Martínez-Sánchez et al., 2010) to avoid the loss due to genetic erosion as a consequence of disasters caused by factors adverse effects such as demographic changes, droughts, destruction of natural areas, substitution of landraces, pests and diseases, among others (CONABIO, 2011; Toledo-Aguilar et al., 2016).
In this sense, it is necessary to rescue and characterize (IBPGR, 1983; UPOV, 2006) the genetic diversity cultivated with the purpose of generating information on the variability of this species that prevails in different regions and agroecological zones of the state of Michoacán. It is also important to explore alternatives that allow us to know and conserve important characteristics to lay the foundations for the genetic improvement of the species.
In the present research work the collection of perón chili morphotypes in different agroecological zones of the state of Michoacán was proposed in order to evaluate the existing morphological variability, as well as the identification of important morphological characteristics to be used in breeding programs of the species.
Materials and methods
The research was carried out in the Phytogenetic Resources Laboratory (LAREFI) of the Advanced Research Unit in Agrobiotechnology (UIIA) of the ‘Presidente Juárez’ School of Agrobiology, located in the city of Uruapan, Michoacán, Mexico. The germplasm collection was carried out through field trips made from February 2016 to August 2017, due to the association of C. pubescens with avocado cultivation, mainly in the area known as the avocado producer fringe. Michoacán, located in the physiographic province of the transversal volcanic system between the parallels 18° 45’ and 20° 6’ north latitude and the meridians 101° 47’ and 103° 13’ west longitude, where ten homogeneous agro-ecological zones predominate (Gutierréz-Contreras et al., 2010).
In total, five fruits and five leaves of 31 cultivated varieties were taken at random. The characterization of fruit, leaf and seed was made based on the morphological descriptors proposed for Capsicum spp. (CATIE-IPGRI-AVRDC, 1995, UPOV, 2006). In total, 27 variables were considered, six of leaf: color, shape, margin of leaf blade, pubescence, length and width; 16 of fruit: volume, presence of anthocyanin stains or stripes, color, shape, length, width, weight, length of the fruit pedicel, thickness of the wall, shape of the fruit in union with the pedicel, neck at the base of the fruit , shape of apex of fruit, appendix in the fruit, transverse wrinkling, number of locules and type of epidermis and five of seed: color, texture, size, weight and number of seeds per fruit.
The experimental design used was completely randomized with five repetitions. For the analysis of qualitative variables, double-state and multi-state data were given to which nominal values were assigned with logical sequence and quantitative variables were standardized to whole numbers with two decimals except for the number of locules and number of seeds per fruit. Subsequently, to determine the existence or not of statistical differences between the characters evaluated, a one-way analysis of variance was performed with the Proc Anova procedure and to evaluate significant statistical differences between cultivated varieties, the Tukey test was used (α≤ 0.05).
In order to select the variables that provided the most information to explain the total variance among the cultivated varieties, 24 variables were selected that presented significant statistical differences, on this list a discriminant analysis of Stepwise was practiced (Romano and Wolf, 2005). Subsequently, to eliminate the correlated variables and avoid problems of collinearity in the matrices, a multiple correlation analysis was made (Pearson and Filon, 1898).
In order to determine the similarity between the cultivated varieties, multivariate statistics tools were used, such as principal component analysis (PCA) and cluster analysis (AC) with the Ward grouping method, with the pseudo F test was determined on degree of reliability of the groups.
For the analyzes described, the SAS computation program version 9.2 was used (SAS, 2012). Finally, to determine the possible relationship of the morphological characteristics with the origin of the cultivated varieties, the georeferencing information of the origin of the cultivated varieties was transformed (FCC, 2018) to construct a matrix of geographical distances (DG), which was correlated with the matrix of Euclidean distances (DE) of morphological characteristics, by means of the Mantel test with the Genalex statistical package (Peakall and Smouse, 2012).
Results and discussion
In four homogeneous agroecological zones (Gutiérrez-Contreras et al., 2010) of the Michoacán avocado producer fringe, it was possible to identify perón chili, in total, 31 cultivated varieties of perón chili with contrasting morphological characteristics. The cultivated varieties were located between 1 623 and 2 282 meters above sea level, height ranges that coincide with those reported for the cultivation of this species (Yáñez et al., 2015; Espinosa-Torres and Ramírez-Abarca, 2016; Caballero-Gutiérrez et al., 2017).
Perón chili is identified as a species that has genetic variability that can be used in breeding programs (Arias et al., 2017). In this study, the cultivated varieties of perón chili showed significant statistical differences (α< 0.05) for 20 variables. In Table 1, simple statistics are presented for each of the variables analyzed. The coefficients of determination (R2) obtained ranged from R2= 0.29 (texture of the seed) to R2= 0.82 (fruit length), indicating that the data fit the model used for the analysis of the information, in addition, most of the variables presented high predictive capacity of the variance.
Table 1. Simple statistics for 27 variables evaluated in 31 cultivated varieties of C. pubescens from the state of Michoacán, Mexico.
Variable | R2* | CV1 | DE2 | Mean | Pr> F3 |
Leaf color | 0.54 | 17.52 | 0.58 | 3.32 | 0.0001 |
Leaf shape | 0.51 | 20.05 | 0.52 | 2.58 | 0.0001 |
Leaf blade margin | 0.39 | 29.26 | 0.5 | 1.7 | 0.0001 |
Leaf pubescence | 0.49 | 16.82 | 0.39 | 2.29 | 0.0001 |
Leaf length | 0.72 | 15.96 | 2.23 | 13.96 | 0.0001 |
Leaf width | 0.64 | 17.85 | 0.91 | 5.08 | 0.0001 |
Fruit volume | 0.75 | 20.55 | 9.12 | 44.39 | 0.0001 |
Spots or streaks | 0.37 | 91.68 | 0.44 | 0.48 | 0.0004 |
Fruit color | 0.73 | 27.95 | 1.09 | 3.9 | 0.0001 |
Fruit shape | 0.3 | 18.33 | 0.82 | 4.46 | 0.015 |
Fruit length | 0.82 | 8.81 | 4.58 | 52.01 | 0.0001 |
Fruit width | 0.81 | 9.25 | 4.66 | 50.34 | 0.0001 |
Fruit weight | 0.81 | 19.75 | 8.17 | 41.38 | 0.0001 |
Pedicel length | 0.57 | 15.42 | 4.63 | 30.02 | 0.0001 |
Wall thickness | 0.36 | 24.32 | 1.35 | 5.56 | 0.0005 |
Fruit shape at the junction with the pedicel | 0.63 | 13.58 | 0.51 | 3.74 | 0.0001 |
Neck at the base of the fruit | 0.56 | 61.27 | 0.36 | 0.59 | 0.0001 |
Fruit apex shape | 0.39 | 13.18 | 0.37 | 2.79 | 0.0001 |
Fruit appendix, vestige of flowering | 0.49 | 95.3 | 0.39 | 0.41 | 0.0001 |
Cross wrinkle | 0.55 | 28.6 | 0.54 | 1.88 | 0.0001 |
Number of locules | 0.32 | 21.69 | 0.54 | 2.48 | 0.007 |
Epidermis type | 0.37 | 28.57 | 0.33 | 1.14 | 0.0004 |
Seed color | 0.34 | 23.26 | 0.53 | 2.26 | 0.002 |
Seed texture | 0.29 | 25.23 | 0.51 | 2.03 | 0.0207 |
Seed size | 0.4 | 27.49 | 0.5 | 1.81 | 0.0001 |
Seed weight per fruit | 0.49 | 28.98 | 0.33 | 1.14 | 0.0001 |
Number of seeds | 0.4 | 26.91 | 17.31 | 64.33 | 0.0001 |
*= coefficient of determination; 1= coefficient of variation; 2= standard deviation; 3= statistical probability of F.
The standard deviation (SD) in most of the collections was low, except for the variables: fruit weight (SD= 8.17), fruit volume (SD= 9.12) and number of seeds per fruit (SD= 17.31). On the other hand, the variability of the evaluated characteristics and interpreted by the coefficient of variation (CV) showed that the measured characters have different levels of dispersion, the characters with the greater variation between collections were: appendix of fruit, vestige of flowering (CV= 95.30) and presence or absence of anthocyanin spots or streaks (CV= 91.68), in contrast to length (CV= 8.81) and fruit width (CV= 9.25).
Similar results have been obtained when characterizing populations of C. annuum, C. baccatum, C. chínense and C. frutescens (Palacios and García, 2009). In this sense, when analyzing collections of C. annuum, C. frutescens and C. chinense CV greater than 25% were obtained for 75% of the evaluated descriptors (Villota-Ceron et al., 2012), in collections of C. annum (timpinchili) the highest CV values were determined for stem length and crown diameter with a CV greater than 50% (Alonso et al., 2012).
The wide variability of morphological characters presented by the genus Capsicum has been observed in different works (Pardey et al., 2009). Although different species belonging to the genus Capsicum share traits in common, each species has its own characteristics, of importance in the selection processes (Palacios and García, 2009). However, the variables evaluated in fruit are mainly considered to discriminate useful genetic variability in breeding programs (Pardey et al., 2006).
For example, the thickness of pulp, weight, length and volume of fruit have been reported as important characteristics in the explanation of the variation that exists within the genus Capsicum (Medina et al., 2006; Castañon-Najera et al., 2008; Bozokalfa et al., 2009; Singh et al., 2017). The results obtained from the stepwise discriminant analysis indicate that the variables that do not provide information on the variability between the cultivated varieties are the volume of fruit and type of epidermis.
Additionally, the results obtained from the stepwise discriminant analysis indicated that the variables: volume and thickness of the fruit wall, do not provide useful information to explain the variation between the cultivated varieties of manzano chili. Likewise, when reviewing the Pearson correlation values, these variables showed a high correlation with the weight and width of the fruit.
In Table 2, there are 27 significant correlations (r> 0.05) identified among the 27 variables evaluated in leaf, fruit and manzano chili seed. Of the 27 correlations, five correlations were identified with values of r≥0.8, the fruit weight with volume and width of fruit (r= 0.9521 and r= 0.8383, respectively), width of fruit, with volume of fruit (r= 0.8197), number of seeds with the weight of seeds (r= 0.8044) and width of the leaf with leaf length (r= 0.7841). 20 correlations with value r≤ 0.5.
In contrast, two correlations with negative value, length of fruit with shape of fruit apex (r= -0.3108) and fruit color with fruit width (r= -0.3114) were identified. Similarly, variables with high values of correlation between leaf and fruit characteristics in collections of the genus Capsicum have been identified (Villota-Ceron et al., 2012).
Table 2. Pearson correlation coefficients identified among the variables evaluated in 31 cultivated varieties of C. pubescens from the state of Michoacán, Mexico.
Variable 1 | Variable 2 | Correlation coefficient | Variable 1 | Variable 2 | Correlation coefficient |
VF | PF | 0.9521 | LF | CBF | 0.3871 |
ACHF | PF | 0.8383 | ACHF | FFUP | 0.3811 |
VF | ACHF | 0.8197 | LF | ARRT | 0.3765 |
PS | NS | 0.8044 | PF | EP | 0.3739 |
LH | ACHH | 0.7841 | VF | FFUP | 0.3732 |
PF | PS | 0.4894 | ACHF | EP | 0.3528 |
LF | LH | 0.4868 | VF | EP | 0.3503 |
VF | PS | 0.4588 | FAF | NS | 0.3205 |
PF | NS | 0.4473 | NL | NS | 0.3182 |
VF | NS | 0.4146 | NL | PS | 0.3175 |
ACHF | PS | 0.4128 | LF | CH | 0.3116 |
LF | LP | 0.4097 | LF | FAF | -0.3108 |
PF | FFUP | 0.4091 | CF | ACHF | -0.3114 |
ACHF | NS | 0.3919 |
Fruit volume (VOL); fruit weight (PF); fruit width (ACHF); seed weight (PS); leaf length (LH); fruit length (LF); fruit apex shape (FAF); number of locules (NL); fruit color (CF); number of seeds (NS); wall thickness (EP); fruit shape at the junction with the pedicel (FFUP); transverse wrinkle (ARRT); leaf width (ACHH); seed color (CS); leaf color (CH); pedicel length (LP); and neck at the base of the fruit (CBF).
In hybrids of C. pubescens the close association of the variables height of the plant, length and width of the leaf is demonstrated, in addition, the effect on the bearing and vigor of the plant as well as on the size of the fruit is pointed out (Arias et al., 2017). The weight of the fruit is a dominant characteristic, related to the size of the fruit and the thickness of the pericarp; these characteristics are important due to consumer demand, since the latter is attracted by large and showy fruits, and fruits with thicker pericarp are more resistant in post-harvest handling (Lannes et al., 2007).
However, the susceptibility to fruit rot caused by Phytophthora was correlated with the thickness of the pericarp; that is, chili with thicker pericarp tend to be more susceptible to P. capsici for this reason, small fruits are considered in breeding programs for their characteristics of resistance to insects and diseases (Naegele et al., 2016) . The results obtained from the analysis of principal components (CP) indicated that the total variance explained, with the first three components was only 45.89%, while with nine, 80% is explained.
Similar results have been obtained in works of morphological characterization in situ in the genus Capsicum, since it has been observed that in this type of work it is not possible to explain more 80% of the variance with the first three components (Castañon-Najera et al., 2008; Martínez-Sánchez et al., 2010; Alonso et al., 2012; Villota-Ceron et al., 2012; de la Cruz-Lázaro et al., 2017). Similar results in this type of work in other species have been reported (Medina-Torres et al., 2015, 2016; Toledo-Aguilar et al., 2016).
In the Figure 1 shows the distribution of 31 cultivated varieties from four homogenous agro-ecological zones based on three main components. When analyzing the eigenvectors with values above 0.3, it is determined that the CP1 is a function of the presence of the neck at the base of the fruit (CBF), leaf length (LH) and fruit length (LF), the CP2 is determined according to the weight of the seed (PS), fruit volume (VOL), width (ACHF) and fruit weight (FP), finally, CP3 is a function of fruit color (CF), transverse wrinkling (ARRT) ), number of locules (NL) and seed color (CS).
Similar results have been reported in others in other species, for example, in C. annuum 74% of the total variance is indicated, explained with the first three main components (Martínez-Sánchez et al., 2010) as a function of days to flowering and fruiting, pericarp thickness, corolla length and width. The most important features to explain the variability between cultivated varieties of different species of the Capsicum genus are those related to the fruit and the architecture of the plant (Castañon-Najera et al., 2008; de la Cruz-Lázaro et al., 2017), these results coincide with those obtained in the present investigation, since the width and length of leaf, shape, length, width and fruit weight have also been identified among the main morphological characteristics that favor the explanation of variability between collections.
Figure 1. Dispersion of cultivated varieties of perón chili from four homogeneous agro-ecological zones of the state of Michoacán; through, of the first three components. Variables associated with CP1= neck in the fruit base (CBF); leaf length (LH) and fruit (LF); variables associated to CP2= seed weight (PS); volume (VOL); width (ACHF) and fruit weight (PF); variables associated with CP3= fruit color (CF); transverse wrinkling (ARRT); number of locules (NL); and seed color (CS).
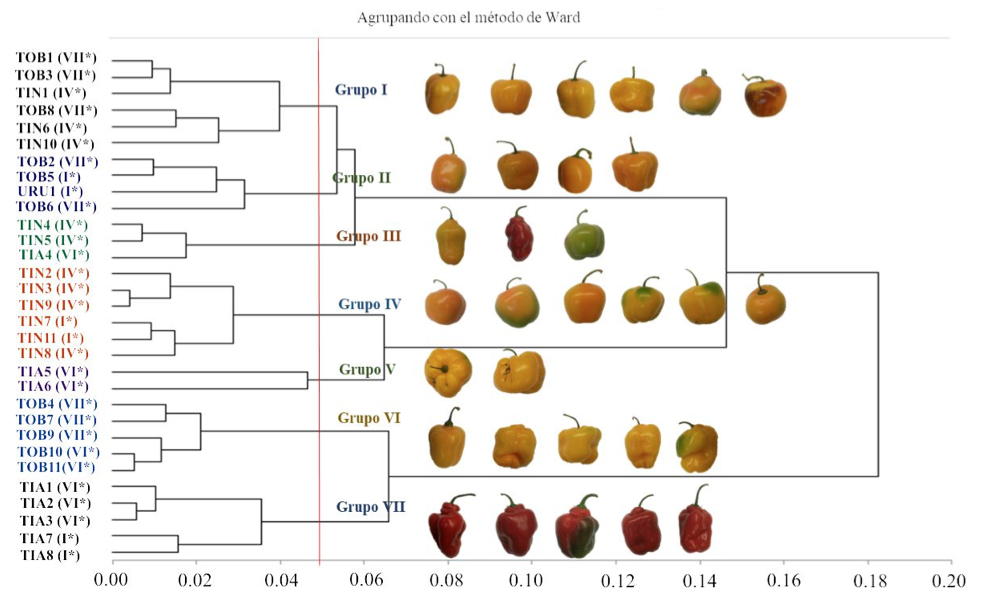
In the dendogram generated from the cluster analysis, groups of cultivated varieties from the same locality were identified, as well as the association of varieties cultivated by the agroecological zone of the place of origin (Figure 2). In the germplasm collected there is a wide variability, these differences may be related to the places of origin. It has been identified that the temperature and the quantity of water available during the growth and reproduction of the plants are important factors for the differentiation of the populations of wild chili that grows in natural conditions (Hernández-Verdugo et al., 2012).
Figure 2. Grouping of 31 cultivated varieties of C. pubescens from four homogeneous agroecological zones* of the state of Michoacán; Toreo el bajo (TOB); Tingambato (TIN); Tiamba (TIA); and Uruapan (URU).
In contrast, the grouping of materials shows no relationship between species or collection environments (Villota-Ceron et al., 2012). The results obtained on the variability and its relation with the origin of the cultivated varieties coincide partially with those indicated by the cited authors, in this work, the existence of groups of cultivated varieties clearly related to the agroecological zones was determined, although they were also identified groups without any apparent relationship to their place of origin. The results obtained in this study corroborate that the existing variability in C. pubescens is mainly explained by characteristics evaluated in the fruits (Table 3).
Table 3. Description of groups of cultivated varieties of C. pubescens based on nine variables that provide the most information to explain the existing total variance.
G1 | MOR2 | CF3 | FF4 | AF5 (mm) | PF6 (g) | FFUP7 | CBF8 | ATF9 | NS10 |
1 | present | yellow to orange | bell-shaped and bell-shaped in block | 46-48 | 34-42 | truncated, cordate and lobed | present and absent | slightly corrugated to intermediate | 50-79 |
2 | present and absent | orange | bell-shaped and bell-shaped in block | 35-46 | 21-33 | truncated | absent | slightly corrugated | 46-73 |
3 | absent | yellow orange and red | almost round, elongated and bell-shaped in block | 36-54 | 21-43 | cordate | present | slightly corrugated | 45-61 |
4 | absent | yellow to orange | almost round | 51-61 | 42-62 | lobed | absent | slightly corrugated | 60-78 |
5 | present and absent | yellow | bell-shaped in block | 63-77 | 60-100 | lobed | absent | intermediate to very corrugated | 88-100 |
6 | present | yellow to orange | bell-shaped and bell-shaped in block | 45-65 | 34-43 | truncated and cordate | present | intermediate to very corrugated | 56-73 |
7 | present and absent | red | triangular and bell-shaped | 40-49 | 25-45 | cordate | present | intermediate to very corrugated | 52-78 |
1= group; 2= anthocyanin stains or streaks; 3= color of fruit; 4= width of fruit; 5= fruit weight; 6= form of the fruit at the junction with the pedicel; 7= neck at the base of the fruit; 8= transverse wrinkling; 9= number of seeds per fruit.
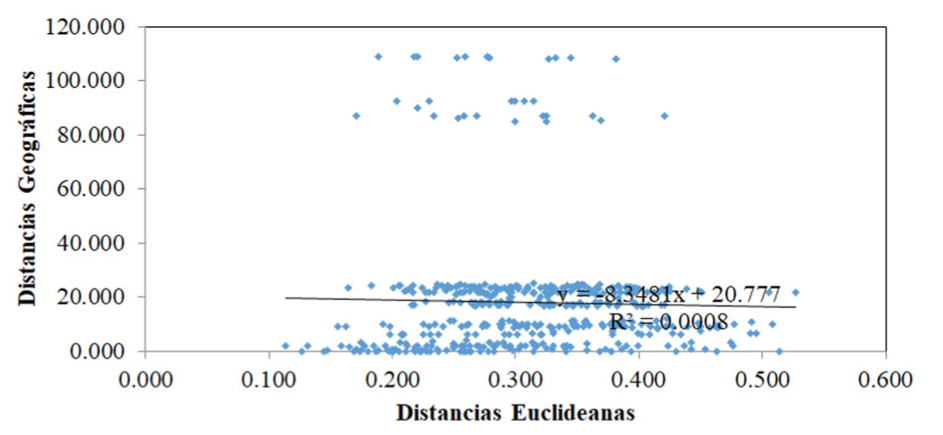
Finally, the results obtained from the Mantel test indicate that there were no significant correlations between the origin of the cultivated varieties and the morphological characteristics evaluated (Figure 3); nevertheless, the data used to correlate the variability with the origin are: latitude north and longitude west, without taking into account the agroecological characteristics of the sites of origin. Therefore, the variability identified among cultivated varieties of C. pubescens is not directly related to geographic location but may be influenced by homogeneous agroecological zones of provenance.
Figure 3. Mantel test, linear correlation between distribution of 31 cultivated varieties of C. pubescens based on Euclidean distances generated by morphological characteristics and geographical distribution according to the geographic coordinates of origin.
In species of the Capsicum genus, the variability within and between species is not necessarily related to the zones of origin (Bobadilla-Larios et al., 2017). Therefore, the grouping of the varieties cultivated in the present study may be influenced by gene flow since the appearance of new forms, new genotypes or hybrids is mainly due to the exchange of germplasm among farmers (Pérez et al., 2015).
Conclusions
The cultivated varieties of perón chili from four homogeneous agro-ecological zones of the state of Michoacán presented a wide morphological variability in leaf tissue, fruit and seed. The characteristics evaluated in the fruit stood out as the most important variables to explain the total variation identified, in addition, that these characteristics can be used as parameters of fruit quality of perón chili.
On the other hand, it was not possible to establish any relationship between the morphological variability and the sites of origin of the cultivated varieties of perón chili evaluated. However, the idea that the morphological characteristics are influenced by the agro-ecological zones of the site of origin and by the gene flow due to the exchange of germplasm among farmers is not discarded.
Acknowledgments
To the CIC-UMSNH, UIAA-Faculty of Agrobiology-UMSNH for the financing of the project and support with the facilities for the execution of the research activities. To the Institutional Program of Doctorate in Agricultural Sciences of the Autonomous University of Nayarit and to the CONACYT by the grant granted for the studies of doctorate.
Cited literature
Alonso, B. R. A.; Zambrano, C. B.; Quiroga, M. R.; Rosales, E. M. A.; y Ponce, D. P. 2012. Caracterización morfológica y molecular de la variabilidad genética del timpinchile (Capsicum annum L. var. Glabriusculum sin. Aviculare) en Chiapas. Rev. Quehacer Científico en Chiapas. 1(13):4-18.
Arias, B. A.; Carranza, J. M.; Estrada, I. M.; Rubí, M. y Vázquez, L. M. 2017. Caracterización morfológica de híbridos de chile manzano. Rev. Mex. Cienc. Agríc. 8(4):825-836.
Arreguin, J.; Bautista, S.; Pérez, M. C. I. y Ramos, M. 2015. Extracción en proceso continuo de oleorresinas de chile seco (Capsicum annuum spp.) a nivel laboratorio. Rev. Cienc. Nat. Agrop. 2(2):200-203.
Barrios-Puente, G.; Espinosa-Torres, L. E.; Figueroa-Hernández, E. y Ramírez-Abarca, O. 2014. Evaluación técnica, financiera y comercial de los sistemas de producción de chile manzano (Capsicum pubescens R. Y P.) en México. Análisis Económico. 29(71):209-219.
Bobadilla-Larios, V.; Esparza-Ibarra, E.; Delgadillo-Ruiz, L.; Gallegos-Flores, P. y Ayala-Lujan, J. L. 2017. Variedades de chile (Capsicum annuum L.) identificadas mediante marcadores RAPD. Trop. Subtrop. Agroecos. 20(3):465-473.
Bozokalfa, M. K.; Esiyok, D. and Turhan, K. 2009. Patterns of phenotypic variation in a germplasm collection of pepper (Capsicum annuum L.) from Turkey. Spanish J. Agric. Res. 7(1):83-95.
Caballero-Gutiérrez, L. B.; Cardozo-Márquez, J. C. y Rojano, B. A. 2017. Effect of the freeze - drying on the functional properties rocoto chilipeper (Capsicum pubescens). Rev. UDCA. Actualidad Divulgación Científica. 20(1):111-119.
Castañon-Najera, G.; Latournerie-Moreno, L.; Mendoza-Elos, M.; Vargas-López, A. y Cárdenas-Morales, H. 2008. Colección y caracterización de Chile (Capsicum spp.) en Tabasco, México. Rev. Inter. Bot. Exp. 77(1):189-202.
CATIE-IPGRI-AVRDC. 1995. Descriptores para Capsicum (Capsicum spp.). Italy, Taiwan y Costa Rica. 110 p.
CONABIO. 2011. Comisión Nacional para el Conocimiento y uso de la Biodiversidad. La Biodiversidad en Puebla: estudio de Estado. México. Gobierno del estado de Puebla-BUAP. (Eds.). Primera edición. 440 p.
de la Cruz-Lázaro, E.; Márquez-Quiroz, C.; Osorio-Osorio, R.; Preciado-Rangel, P. y Márquez-Hernández, C. 2017. Caracterización morfológica in situ de chile silvestre pico de paloma (Capsicum frutescens L.) en Tabasco, Mexico. Acta Universitaria. 27(2):10-16.
Espinosa-Torres, L. E. y Ramírez-Abarca, O. 2016. Rentabilidad de chile manzano (Capsicum pubescens R. Y P. ) producido en invernadero en Texcoco, Estado de México. Rev. Mex. Cienc. Agríc. 7(2):325-335.
FCC. 2018. Degrees minutes seconds to/from decimal degrees. Federal Communications Commission. https://www.fcc.gov/media/radio/dms-decimal.
Gutiérrez-Contreras, M.; Lara-Chávez, B. N.; Guillén-Andrade, H. y Chávez-Bárcenas, A. T. 2010. Agroecología de la franja aguacatera en Michoacán, México. Interciencia. 9(35):647-653.
Hernández-Verdugo, S.; Porras, F.; Pacheco -Olvera, A.; López-España, R. G.; Villarreal-Romero, M.; Parra-Terraza, S. y Osuna-Enciso, T. 2012. Caracterización y variación ecográfica de poblasiones de chile (Capsicum annum var. Glabriusculum) silvestre del noroeste de México. Polibotánica. 33(1):175-191.
Hernández-Verdugo, S. 2014. Importancia del chile silvestre (Capsicum annuum) como recurso genético de México. Mensaje Bioquímico. 41(XLI):289-304.
IBPGR. 1983. Crop genetic resourses centre plant production and protection división Food and Agriculture Organization of the United Nations Via delle Terme di Caracalla. Rome, Italy. Italy. ISBN 92-9043-108-3. 451p.
Lannes, S. D.; Finger, F. L.; Schuelter, A. R. and Casali, V. W. D. 2007. Growth and quality of Brazilian accessions of Capsicum chinense fruits. Sci. Hortic. 112(3):266-270.
López-López, Á.; Espinoza-Santana, S.; Ceceña-Duran, C.; Ruiz-Alvarado, C.; Núñez-Ramírez, F. y Araiza-Zúñiga, D. 2015. Biosintesis, acumulación y efecto del ambiente sobre compuestos antioxidantes del fruto del cultivo del chile (Capsicum spp.). Rev. Cienc. Biol. Salud. 17(2):50-57.
Martínez-Sánchez, D.; Pérez-Grajales, M.; Rodríguez-Pérez, J. E.; Del, E. y Moreno-Pérez, C. 2010. Colecta y caracterización morfológica de chile de agua (Capsicum annuum L.) en Oaxaca, México. Rev. Chapingo Ser. Hortic. 16(3):169-176.
Medina, C. I.; Lobo, M. y Farley, G. A. 2006. Variabilidad fenotípica en poblasiones de ají y pimientón de la colección colombiana del género Capsicum. Rev. Corpoica-Ciencia y Tecnología Agropecuaria. 7(2):25-39.
Medina-Torres, R.; Juárez-López, P.; Salazar-García, S.; López-Guzmán, G. G.; Ibarra-Sanchez, L. S.; Arrieta-Ramos, B. G. y Martínez-Moreno, E. 2015. Evaluación de calidad en frutos de 41 genotipos de nanche (Byrsonima crassifolia L. HBK) de Nayarit , México. Revista Mex. Cienc. Agríc. 16(2):253-264.
Medina-Torres, R.; Salazar-García, S.; Ibarra-Estrada, M. E. y López-Guzmán, G. G. 2016. Descriptores morfológicos de hoja y fruto de selecciones comerciales de nanche (Byrsonima crassifolia (L.) HBK) en Nayarit, México. Rev. Bio Ciencias. 3(4):269-285.
Moreno-Pérez, del C. E.; Avendaño-Arrazate, C. H.; Mora-Aguilar, R.; Cadena-Iñiguez, J.; Heber Aguilar-Rincón, V. y Aguirre-Medina, J. F. 2011. Diversidad morfológica en colectas de chile guajillo (Capsicum annuum L.) del Centro-Norte de México. Rev. Chapingo Ser. Hortic. 17(1):23-30.
Naegele, R. P.; Mitchell, J. and Hausbeck, M. K. 2016. Genetic diversity, population structure, and heritability of fruit traits in Capsicum annuum. PLoS ONE. 11(7):1-17.
Palacios Castro, S. y García Dávila, M. A. 2009. Caracterización morfológica de 93 accesiones de Capsicum spp del banco de germoplasma de la Universidad Nacional de Colombiana Sede Palmira. Acta Agron. 57(4):247-252.
Pardey, C. R.; Agr, I.; García, D. M. A. y Vallejo, C. F. A. 2006. Caracterizacion morfologica de cien introducciones de Capsicum de banco de germoplasma de la Universidad Nacional de Colombia sede Palmira. Acta Agron. 55(3):1-10.
Pardey, R. C.; García, D. M. A. y Vallejo, C. F. A. 2009. Evaluación agronómica de accesiones de Capsicum del banco de germoplasma de la Universidad Nacional de Colombia sede Palmira. Acta Agron. 58(1):23-28.
Peakall, R. and Smouse, P. E. 2012. GenALEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics. 28(19):2537-2539.
Pearson, K. and Filon, L. N. G. 1898. VII. Mathematical contributions of the theory of evolution. IV. On the probable errors of the frequency constants and correlation. Philosophical Transactions of the Royal Society of London A. 191:229-311. Doi: 10.1098/rsta.1898.0007.
Pérez, L.; Castañon, G.; Ramírez, M. y Mayek, N. 2015. Avances y perspectivas sobre el estudio del origen y la diversidad genética de Capsicum spp. Ecosistemas y Recursos Agropecuarios. 2(4):117-128.
Romano, J. P. and Wolf, M. 2005. Stepwise multiple testing as formalized data snooping. Econometrica. 73(4):1237-1282.
Sánchez-Sánchez, H.; González-Hernández, V. A.; Cruz-Pérez, A. B.; Pérez-Grajales, M.; Gutiérrez-Espinosa, M. A.; Gardeja-Bejar, A. A. y Gómez-Lim, M. A. 2010. Herencia de capsaicinoides en chile manzano (C. pubescens R. y P.). Agrociencia. 44(6):655-665.
SAS Institute Inc. 2012. Introduction to bayesian analysis procedures. In: SAS/STAT 9.2 User’s Guide, Ch. 7. SAS Institute, Inc., Cary, NC. 141-179 pp.
SIAP. 2019. Servicio de Información Agroalimentaría y Pesquera, México. http://infosiap.siap.gob.mx:8080/agricola-siap-gobmx/AvanceNacionalCultivo.do.
Singh, P.; Jain, P. K. and Sharma, A. 2017. Genetic variability, heritability and genetic advance in chilli (Capsicum annuum L.) genotypes. Inter. J. Current Microbiol. Appl. Sci. 6(9):2704-2709.
Toledo-Aguilar, R.; López-Sánchez, H.; López, P. A.; Rodríguez, J. D. D. G. y Santacruz, A. 2016. Diversidad morfológica de poblaciones nativas de chile poblano. Rev. Mex. Cienc. Agríc. 7(5):1005-1015.
UPOV. 2006. International union for the protection of new varieties of plants. Sweet pepper, hot pepper, paprika, chili. https://doi.org/10.1147/sj.414.0732.
Villota-Ceron, D.; Bonilla-Betancourt, M. L.; Carmen-Carrillo, H.; Jaramillo-Vázquez, J. y García-Davila, M. A. 2012. Caracterización morfológica de introducciones de Capsicum spp. existentes en el banco de germoplasma activo de Corpoica C.I. Palmira, Colombia. Acta Agron. 61(1):16-26.
Yáñez, P.; Balseca, D.; Rivadeneira, L. y Larenas, C. 2015. Características morfológicas y de concentración de capsaicina en cinco especies nativas del género Capsicum cultivadas en Ecuador. La Granja. 22(2):12-32.
Zegbe, D. J. A.; Valdez, C. R. D. y Lara, H. A. 2012. Reseña de libro. Cultivo de chile en México. Rev. Fitotec. Méx. 35(4):264-264.