Revista Mexicana de Ciencias Agrícolas volumen 10 número 8 12 de noviembre - 31 de diciembre, 2019
DOI: https://doi.org/10.29312/remexca.v10i8.1955
Artículo
Caracterización de colectas de chía de la región occidental de México
Lucero Carolina Medina-Santos
Jorge Covarrubias-Prieto
César L. Aguirre-Mancilla
Gabriel Iturriaga
J. Gabriel Ramírez-Pimentel
Juan Carlos Raya-Pérez§
Tecnológico Nacional de México-Instituto Tecnológico de Roque. Carretera Celaya-Juventino Rosas km 8, Roque, Celaya, Guanajuato, México. CP. 38110. (ncarolinanm@hotmail.com; jor-covarru-jrg@hotmail.com; ceaguirre@itroque.edu.mx; gaiturriaga@itroque.edu.mx: garamirez@itroque.edu.mx).
§Autor para correspondencia: juraya@itroque.edu.mx.
Resumen
La chía es un recurso muy valioso ya que es la principal fuente de ácidos grasos insaturados, principalmente α-linolénico, además de proteína y fibra. En esta investigación se realizaron siete colectas en siete localidades para su estudio y comparación. Se midió tamaño de semilla, peso de mil semillas, peso volumétrico, color, porcentaje de germinación y extracción de aceite para análisis en cromatografía. La colecta de chía cultivada de Guanajuato presentó mayor variabilidad en tamaño de semilla y la de Roque la menor. Las colectas silvestres tuvieron el menor tamaño de semilla. La cultivada de Guanajuato tiene el peso mayor en semilla y las silvestres el menor, estas últimas no germinaron, pero sí lo hicieron las domesticadas. No hubo diferencias en peso volumétrico; sin embargo, si hubo diferencias en los cromatogramas (HPLC) y en las características físicas y variación de color en las domesticadas, incluidas semillas de color blanco. Esta investigación prospectiva muestra que la variabilidad presente en la chía silvestre y cultivada puede ser aprovechada para el fitomejoramiento.
Palabras clave: Hirsuta, Salvia hispanica, S. tiliifolia, ácido graso, diversidad.
Recibido: septiembre de 2019
Aceptado: noviembre de 2019
Introducción
La domesticación es el resultado de un proceso de selección que lleva a las plantas a una mejor adaptación al manejo humano, aunque estas adaptaciones puedan comprometer su supervivencia en la naturaleza (Brown 2010). En México, a pesar de ser el centro de domesticación de la chía (Salvia hispanica), hay pocos estudios de variabilidad genética; en 2008 se realizó una caracterización morfológica de 22 muestras de chía recolectada en distintos sitios de centro y Sudamérica, así como varios estados de México incluyendo a: Sonora, Chihuahua, Jalisco, Morelos y Guerrero (Palma-Rojas et al., 2017).
Se contemplaron variedades silvestres y domesticadas, la mayor parte de estas muestras tuvieron diferencias morfológicas muy marcadas. Se sabe que existen poblaciones silvestres a lo largo de todo el país, sin embargo, no hay indicadores fuera de su origen geográfico que ayuden a diferenciarlas y las propiedades bioquímicas de la mayoría de estas variedades son desconocidas. Es posible que al no tener esta información se estén desperdiciando genotipos productivos importantes en el mejoramiento genético de la chía; ya que la variabilidad es esencial para la sustentabilidad (Brown, 2010).
La chía es una especie diagnóstica que define las fronteras de Mesoamérica (Cahill, 2004), es un cultivo anual que se distribuye en ambientes semicálidos y templados, se adapta a suelos arcillosos y arenosos que estén bien drenados, sin embargo, requiere niveles adecuados de nitrógeno para proporcionar buenos rendimientos (Herman et al., 2016). Es una planta de día corto, no tolera las heladas y requiere abundante sol para producir; en países del primer mundo se han generado variedades de día largo y capaces de producir en distintas condiciones climáticas (Jamboonsri et al., 2012; Grimes et al., 2018). Actualmente la chía se cultiva en varios países de Latinoamérica obteniendo rendimientos de hasta 3 t ha-1 (Baginsky et al., 2016; Muñoz Máximo et al., 2017).
En las últimas décadas ha resurgido, las propiedades nutraceúticas y sus atractivos beneficios nutricionales han ampliado su consumo (Xingu et al., 2017). La distribución de este cultivo crece aceleradamente, sin embargo, existen pocas fuentes de información que puedan proporcionar datos actualizados respecto a su producción y distribución mundial (Jamboonsri et al., 2012; Baginsky et al., 2016; Grimes et al., 2018; Win et al., 2018), S. hispanica y S. tiliifolia son las dos especies que se utilizan para la producción de semilla.
La utilidad de la recolección y estudio de los recursos genéticos se ejemplifica en el jitomate o tomate (S. Lycopersicum) en el que se han investigado la morfología, fisiología y el rendimiento, tanto de la especie cultivada como de sus parientes silvestres (Ichihashi y Sinha, 2014). La semilla de chía contiene 32% de aceite y 20% de proteína; contiene aminoácidos esenciales como lisina y aminoácidos azufrados (metionina y cisteína). Tiene alto contenido de antioxidantes (Oliveira-Alves et al., 2017), y posee compuestos con actividad biológica en diferentes partes de la planta (Marineli et al., 2015; Fan et al., 2019; Kobus-Cisowska et al., 2019; da Silva, 2015).
La chía es la planta con mayor contenido de ácido graso α-linolénico, precursor del eicosapentaenoico y docosahexaenoico, ácidos grasos importantes para el desarrollo de la cognición, la visión y que reducen el riesgo de enfermedades coronarias. La semilla contiene además 40% de fibra dietética y calcio (Sreedhar et al., 2015; Medina-Santos et al., 2017). La localidad de siembra tiene efecto sobre el contenido de α-linolénico, linoeico y oleico y sobre la relación omega6/omega3 (Ayerza y Coates, 2009a). El objetivo de esta investigación fue caracterizar y comparar colectas de chía silvestre (S. tiliifolia y S. hirsutum) y cultivada (S. hispanica) a fin de contribuir al conocimiento de la diversidad de este importante recurso fitogenético.
Materiales y métodos
Se realizaron colectas de cultivares de chía, espigas en madurez fisiológica, en siete localidades: 1) Roque (20° 35’ 00.00’’ latitud norte 100° 50’ 11.54’’ longitud oeste), Celaya, Guanajuato (silvestre y domesticado); 2) Guanajuato (21° 00’ 02.88’’ latitud norte 101° 14’ 24.56’’ longitud oeste), Guanajuato (domesticado); 3) Ballesteros, Salvatierra (20° 13’ 33.99’’ latitud norte 100° 48’ 38.42’’ longitud oeste), Guanajuato (silvestre); 4) Comala (19° 19’ 09.09’’ latitud norte 103° 46’ 18.08’’ longitud oeste), Colima (domesticado); 5) Irapuato (20° 16’ 09.61’’ latitud norte 10° 13’ 448.31’’ longitud oeste), Guanajuato (silvestre y domesticado); 6) Uruapan (19° 26’ 34.09’’ latitud norte 10° 20’ 141.01’’ longitud oeste), Michoacán (silvestre); 7) La Calera, Sixto Verduzco (20° 16’ 09.61’’ latitud norte 101° 34’ 48.31’’ longitud oeste), Michoacán (silvestre).
Las domesticadas corresponden a la especie S. hispanica y la de La Calera e Irapuato a la especie S. tiliifolia; la de Salvatierra a la especie S. hirsuta. (Calderón de Rzedoswski y Rzedowski, 2004). Para obtener la semilla las inflorescencias se sometieron a secado a temperatura ambiente y posteriormente se sacudieron con el fin de liberar las semillas, que fueron guardadas en bolsas plásticas con su respectiva identificación a temperatura ambiente en un lugar fresco y seco.
Para medir las semillas, estas fueron fotografiadas sobre papel milimétrico y posteriormente se cuantificaron los pixeles por medio del software de medición Golden Ratio, la longitud y el grosor de las semillas en milímetros se calcularon por medio de la siguiente fórmula.
Para determinar el peso de semilla se realizaron tres pruebas: peso individual de semilla, peso de 1 000 semillas y peso volumétrico. El peso individual se obtuvo pesando 50 semillas por medio de una balanza analítica. Para la determinación de peso de 1000 semillas se realizaron 4 repeticiones de 200 semillas, se pesaron y se obtuvo el promedio, este promedio fue multiplicado por 5 de acuerdo al manual de la ISTA (ISTA, 2005).
El peso volumétrico se realizó pesando la semilla que cabe en un recipiente de volumen conocido, el cual se llenó dejando caer la semilla de un embudo desde una distancia aproximada de 5 cm sobre la parte central del recipiente para una distribución uniforme hasta el llenado del recipiente, el exceso de semilla de eliminó pasando una regla por el borde de este. Después de obtener el peso, se realizó la conversión a kg hL-1.
La identificación del color de semilla se realizó al tomar muestras de 50 semillas de cada cultivar que fueron inspeccionadas visualmente y fotografiadas para conocer la uniformidad del color, en los cultivares en los que se presentó más de un color de semilla, estas fueron contadas y se determinó el porcentaje de semillas de cada color. Para obtener el porcentaje de germinación se realizaron tres repeticiones de 100 semillas en papel de germinación húmedo a una temperatura de 25 °C por 8 días en ausencia de luz, al terminar este periodo, se realizó el conteo de plántulas normales, anormales y semillas no germinadas, y se obtuvo un promedio del porcentaje de plántulas normales (ISTA, 2005).
Para la extracción de aceite se utilizó el método de Folch modificado (1975); se muele la muestra e incuba en una solución de cloroformo-metanol 50:50 (v/v) en agitación constante a temperatura ambiente; la relación de la solución con la harina para la maceración fue de 4:1 (v/p). Posterior a la maceración se realizó un prensado manual y un filtrado en 8 capas de tela de gasa. El líquido obtenido en la filtración se dejó reposar a temperatura ambiente de 2-3 días hasta volatizar la mezcla de solventes obteniéndose el aceite en el frasco (Puttini et al., 2005) El aceite fue transesterificado en baño maría a temperatura constante (65-68 ºC), la reacción se realizó por el método propuesto por Torossi (2006).
Para el análisis se utilizó un cromatógrafo (Thermo Scientific) para HPLC equipado con una columna hidrofóbica C18 y un detector de absorbencia de UV. Los esteres de ácidos grasos fueron diluidos en metanol grado HPLC (1:1 000) según lo recomendado en el manual de HPLC de Kromidas (2006), el método utilizado para la separación fue la inyección de metanol-agua con una concentración de metanol inicial 100% y una concentración final 60% Kromidas (2006) Como comparación se usó aceite comercial de chía (extraído y elaborado en Jalisco). Los datos fueron analizados empleando un diseño completamente al azar usando el programa SAS 9.1.
Resultados y discusión
Las accesiones de chía cultivada evaluadas mostraron diferencias altamente significativas en tamaño y peso de semilla (Cuadro 1), lo que permite asumir que presentan diferencias fenotípicas y genotípicas. La media de la longitud de semilla de la colecta de Colima fue de 1.95 mm, la de Guanajuato presentó un rango de 1.67-2.3 con una media de 2 mm. La colecta de Irapuato presentó valores entre 1.69 a 3.32 mm. La colecta de Guanajuato presentó mayor variabilidad en longitud de semilla y la chía de Roque presentó la menor variabilidad en esta característica.
Cuadro 1. Cuadrados medios, grados de libertad y significancia estadística del análisis de varianza de la longitud y peso de semilla en colectas de chía. Celaya, P-V 2016.
FV | GL | Cuadrados medios | |
Long semilla | Peso de semilla | ||
Cultivar | 7 | 5.48726** | 0.7266** |
Error | 32 | 0.01006 | 0.0011 |
Total | 39 | ||
**= indica significancia estadística al nivel 0.01 de probabilidad.
La semilla de chía silvestre de La Calera presentó valores entre 1.26 y 1.47 mm con una media de 1.36 mm. La semilla de la colecta de Irapuato tuvo una media de 1.35 mm. La semilla de la colecta de Roque presentó una media de 1.36 mm. La chía silvestre de Salvatierra tuvo una media de 1.43 mm. En condiciones naturales, la semilla grande presenta mayor porcentaje de germinación y muestra mayor vigor, comparada con las pequeñas.
Tenorio-Galindo et al. (2008) señalan que la variación en tamaño y color de la semilla se puede relacionar con capacidad de germinación bajo distintas condiciones ambientales. Jamboonsri et al. (2012), mencionan que el aumento de tamaño de la parte de interés antropocéntrica es una de las características de las plantas cultivadas, en este caso la semilla de las formas cultivadas presentó el doble de tamaño que las silvestres (Cuadro 2).
La colecta de Roque cultivada tuvo un tamaño de semilla estadísticamente superior al resto de colectas; la semilla silvestre de Roque presentó la mayor absorbencia a los o.4 min, el siguiente grupo respecto a esta variable estuvo conformado por Roque cultivada y la Calera silvestre (Cuadro 2). Las colectas silvestres de La Calera, Roque e Irapuato, como era de esperarse, tuvieron el tamaño de semilla estadísticamente más pequeño. Hoyle et al. (2008) encontraron en Goodenia fascicularis, que cuando crecen en ambiente cálido, aceleran la fase reproductiva y producen más semilla, mientras que las que crecen en ambiente fresco, acumulan más biomasa y producen menos semilla, pero esta semilla es más grande y tiene mayor viabilidad.
Cuadro 2. Comparación de medias mediante la prueba de Tukey de las variables peso individual, longitud de semilla y absorbencia (minuto 0.4) de ocho colectas de chía. Celaya, Guanajuato. P-V 2016.
Cultivar | Peso (mg) | Longitud (mm) | Absorbencia (%) |
C. Guanajuato | 1.285 a | 2 b | 0.696 c |
C. Irapuato | 1.135 b | 1.98 b | 0.731 c |
C. Colima | 1.125 bc | 1.94 c | 0.496 d |
C. Roque | 0.977 c | 2.05 a | 0.752 bc |
S. Salvatierra | 0.475 d | 1.43 d | 0.647 c |
S. Irapuato | 0.43 d | 1.35 e | 0.651 c |
S. Roque | 0.43 d | 1.36 e | 0.876 a |
S. La Calera | 0.406 d | 1.36 e | 0.848 b |
Medias con la misma letra dentro de cada variable son estadísticamente iguales (Tukey, p< 0.05).
Ahmed y Fayyaz-ul-Hassan et al. (2015) encontraron que, en trigo, el estrés hídrico incrementa el contenido de proteína en el grano y reduce el tamaño del mismo. Ayerza y Coates (2009a) mencionan que el contenido de proteína en chía es influenciado por el ambiente. En trigo, el mayor rendimiento se logra con mayor peso de grano, lo que valdría la pena considerar en la selección de cultivares de chía (Sun et al., 2017).
Los bajos rendimientos en chía pueden deberse al bajo peso de la semilla y al bajo contenido de aceite. Ayerza (2010) menciona que a medida que la altitud decrece la temperatura aumenta y el contenido de aceite disminuye, aunque a los 1621 msnm encontró que la relación ω6-ω3 disminuye, lo que es bueno para la salud de los consumidores. Celaya se encuentra a 1750 msnm, muy cercano al valor reportado por Ayerza (2010).
C: cultivada; S: silvestre. En el análisis de varianza del peso volumétrico de las muestras no se encontró diferencia significativa entre los cultivares; se encontraron valores de 78.5 (kg hL-1) para la colecta de silvestre de Irapuato y de 66.17 (kg hL-1) para la colecta cultivada de Irapuato. Vázquez-Carrillo et al. (2012) encontraron que en maíz el tamaño del grano no influyó sobre el peso hectolítrico, pero factores ambientales como la precipitación podría afectarlo. Dado que se trabajó con colectas silvestres no se podría asegurar si las plantas tuvieron suficiente agua, lo mismo que las domesticadas, lo que podría explicar por qué no hubo diferencias en esta característica.
En el Cuadro 3 se muestran los resultados de las pruebas de germinación estándar, se puede observar que las semillas silvestres no presentaron germinación; estas semillas tienen latencia por ser silvestres, ya que en la naturaleza esta característica es necesaria para germinar a diferentes tiempos y cuando las condiciones son óptimas para asegurar la supervivencia de la especie. Las semillas de los cultivares domesticados presentaron porcentajes de germinación de 78%, relativamente bajo comparado con especies altamente domesticadas.
Cuadro 3. Porcentaje de germinación de colectas de chía, silvestres y cultivadas, Celaya P-V 2016.
Cultivar | (%) de germinación semilla sin tratar | (%) germinación semilla tratada con ácido giberélico |
C. Colima | 87 | - |
C. Guanajuato | 80 | - |
C. Irapuato | 71 | - |
C. Roque | 74 | - |
S. La Calera | 0 | 5 |
S. Irapuato | 0 | 7 |
S. Salvatierra | 0 | 2 |
S. Roque | 0 | 9 |
Este resultado está acorde con Lobo et al. (2007) quienes señalan que las semillas pueden presentar latencia morfofisiológica, con embriones no totalmente desarrollados y con mecanismos fisiológicos que inhiben la germinación; la impermeabilidad de la testa podría ser la que puede estar impidiendo la germinación. Los estudios de en canola muestran que la germinación depende del contenido de ácidos grasos y otras reservas; en maíz se halló una correlación positiva entre el contenido de lípidos y el porcentaje de germinación (Seyyedi et al., 2018).
A las semillas silvestres se les puede aplicar un tratamiento con ácido giberélico, ya que este es capaz de inducir la germinación al ayudar a romper la latencia. Con la percepción de giberelinas, muchas enzimas se activan e inician el proceso de degradación del endospermo.
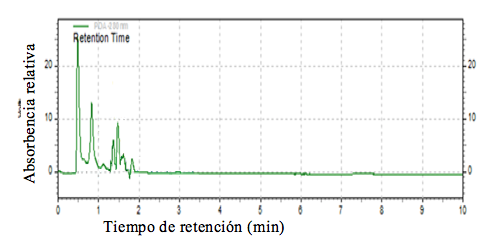
La Figura 1 muestra el cromatograma representativo de la chía domesticada de Colima, en el cual podemos observar la aparición de un pico al minuto 0.828 que no aparece en el cromatograma del aceite comercial. El pico mayor, representa al ácido α-linolénico, sin embargo, el porcentaje de absorbencia es mayor al porcentaje de ácido linolénico en chía comercial reportado por Gutiérrez (2014). La chía contiene entre 5 y 16 ácidos grasos, dependiendo de las condiciones de crecimiento; los más comunes son palmítico, esteárico, oleico, linoleico y α-linolénico. Ayerza y Coates (2009a) no encontraron diferencias en cuatro genotipos y reportaron un contenido de ácido linoleico de 20% y de α-linolenico de 61.7% en la composición del aceite.
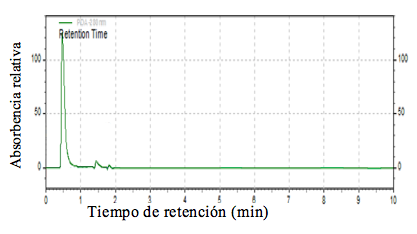
Estos investigadores (2009) reportaron una selección de chía llamada Tliltic que tiene un menor contenido de ácido linoléico (ω6), lo que podría aprovecharse para lograr variedades con una buena relación ω3: ω6. Los picos que se presentan en los diferentes cultivares varían en tamaño y se presentan o no de acuerdo con la composición del aceite de cada cultivar. Se eligieron los picos más destacados que aparecen en las tres repeticiones de cada colecta. Es posible apreciar que en los cromatogramas existen tres picos comunes para todos los materiales; sin embargo, el perfil de aparición de los picos es diferente para todas las colectas (Figuras 1, 2 y 3). En la Figura 2 se muestran los porcentajes de absorbencia de los ésteres del aceite de chía silvestre de La Calera, se puede observar que el porcentaje de absorbencia del primer pico (minuto 0.466) es bastante alto, después de este pico, los más visibles son los del minuto 1.4 y 1.8.
Figura 1. Cromatograma de los picos del aceite de la chía de Colima (cultivada).
Figura 2. Cromatograma de picos de aceite extraído de semilla de chía de Calera (S. tiliifolia, silvestre).
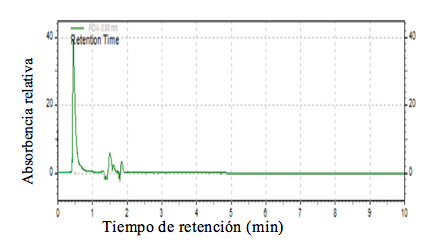
Figura 3. Cromatograma de los picos del aceite extraído de semilla de chía de Salvatierra (S. hirsutum).
Las características generales de las semillas de chía se presentan en los Cuadros 4 y 5 y se muestran fotografías de algunas de ellas (Figuras 4 y 5). Se puede observar la gran similitud entre las semillas de las colectas, todas fueron de forma ovalada y con pericarpio liso.
Figura 4. Porcentaje de colores encontrados en la semilla de chía silvestre de La Calera, Michoacaán.
Figura 5. Porcentaje de colores encontrados en el cultivar de semilla de chía cultivada de Colima.
Cuadro 4. Características de las semillas de chía silvestre.
Características generales de las semillas silvestres | Localidades | |||
Irapuato | La Calera | Roque | Salvatierra | |
Tamaño promedio (mm) | 1.36 | 1.36 | 1.43 | 1.43 |
Peso promedio por semilla (mg) | 1.8 | 1.5 | 1.7 | 1.8 |
Color predominante | Gris con líneas negras | Café con manchas blancas | Gris con líneas negras | Café con manchas blancas |
Cuadro 5. Características de las semillas de chía cultivada.
Características generales de las semillas | Localidades | |||
Irapuato | Colima | Guanajuato | Roque | |
Tamaño (mm) | 2.2 | 2 | 1.9 | 2 |
Peso promedio por semilla (mg) | 4.5 | 4.7 | 5 | 5.2 |
Color predominante | Gris con líneas negras | Café claro con líneas café oscuro | Gris oscuro con líneas negras | Café claro con líneas café oscuro |
Cahill (2004) localizó, al estudiar 348 colectas de chía silvestre y domesticada, mayor diversidad entre las variedades silvestres que entre todas las domesticadas; señala que hay poco entrecruzamiento entre silvestres y domesticadas, con diferencias entre unas y otras a nivel de anteras, cáliz y morfología de la semilla. En las variedades de Centroamérica no encontró gigantismo en la semilla, con respecto a lo hallado en las variedades mexicanas. Jamboonsri et al. (2012) señalan que las plantas domesticadas producen semillas grandes y no presentan mecanismos de dispersión.
Entre las plantas silvestres no se encontraron semillas de color blanco, en las plantas cultivadas se reporta hasta 1% de semillas de color blanco. Ayerza (2010) no encontró diferencias en el contenido de aceite entre las semillas oscuras y las de color blanco, aunque dado que se supone una preferencia de los consumidores por las de color blanco, ya se tienen selecciones de chía con este color de semilla; el color blanco está gobernado por un gen recesivo, llamado scc, lo cual explicaría en parte la ausencia de semillas de este color en las colectas silvestres. En aspecto general, las semillas fueron muy similares entre las colectas, aunque variaron en tamaño (Cuadros 4, 5).
Conclusiones
Las semillas provenientes de las colectas de plantas silvestres tienen un menor tamaño que las producidas por las plantas cultivadas. En peso ocurre lo mismo, las semillas de las plantas domesticadas presentaron más del doble de peso que las semillas de las plantas silvestres. Las domesticadas presentan semillas de color blanco. La chía silvestre, S. tiliifolia e S. hirsuta muestran un pico mayoritario, aproximadamente entre 8 y 10 veces en mayor cantidad que los demás ácidos grasos presentes, que indica un alto contenido de α-linolénico, como la chía domesticada, S. hispanica, muestra el pico, pero es menor y estadísticamente diferente al de la chía silvestre.
Agradecimientos
Al Dr. Jesús G. González Gallegos por la identificación de la especie tiliifolia.
Literatura citada
Ahmed, M. and Fayyaz-ul-Hassan. 2015. Response of spring wheat (Triticum aestivum L.) quality traits and yield to sowing date. Plos One 10(4):e0126097.doi:10.137/journal.pone.0126097.
Ayerza, R. 2010. Effects of seed color and growing locations on fatty acid content and composition of two chía (Salvia hispánica L.) genotypes. J. Am. Oil Chem. Soc. 87(10):1161-1165.
Ayerza, R. and Coates, W. 2009a. Some quality components of four chía (Salvia hispanica L.) genotypes agrown under tropical coastal desert ecosystems. Asian J. Plant Sci. 8(4):301-307.
Ayerza, R. and Coates, W. 2009b. Influence of environment on growing period and yield, protein, oil and α-linolenic content of three chía (Salvia hispanica L.) selections. Industrial Crop Products. 30(2):321-324.
Baginsky, C.; Arenas, J.; Escobar, H.; Garrido, M.; Valero, N.; Tello, D.; Pizarro, L.; Valenzuela, A.; Morale, L. and Silva, H. 2016. Growth and yield of chía (Salvia hispanica L.) in the Mediterranean and desert climates of Chile. Chilean J. Agric. Res. 76(3):255-264.
Brown, A. H. D. 2010. Variation under domestication: 1859 and today. Phil. Trans. R. Soc. B. 365:2523-2530.
Cahill, J. 2004. Genetic diversity among varieties of chía (Salvia hispanica L.). Gen. Res. Crop Evol. 51(7):773-781.
Calderon de Rzedowski, G. y Rzedowski, J. 2004. Manual de malezas de la región de Salvatierra, Guanajuato. Flora del Bajío y regiones adyacentes XX. Instituto de Ecología, Xalapa, Ver. 316 p.
da Silva, R., 2015. Antioxidant potential of dietary chía seed and oil (Salvia hispanica L.) in diet-induced obese rats. Food Res. Intern. 76(3):666-674.
Fan, M.; Luo, D.; Peng, L.; Li, X.; Wu, X.; Ji, X. and Zhao, Q. 2019. Phytochemistry Neo-clerodane diterpenoids from aerial parts of Salvia hispanica L. and their cardioprotective effects. Phytochemistry.166:1-11. doi: 10.1016/j.phytochem.2019.112065.
Folch, J.; Lees, M. and Stanley, G. H. 1957. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 226:497-509.
Grimes, S. J.; Phillips, T. D.; Hahn, V., Capezzone; F., & Graeff-Hönninger; S. 2018. Growth, Yield Performance and Quality Parameters of Three Early Flowering Chia (Salvia Hispanica L.) Genotypes Cultivated in Southwestern Germany. Agriculture. 8(10):154-173.
Gutiérrez, T. R.; Ramírez, V. M. L.; Vega-León, S.; Fontecha, J.; Rodríguez, L. M. y Escobar, M. A. 2014. Contenido de ácidos grasos en semillas de chía (Salvia hispanica L.) cultivadas en 4 estados de México. Rev. Cubana de Plantas Medicinales. 19(3):199-207.
Herman, S.; Marco, G.; Cecilia, B.; Alfonso, V.; Luis, M.; Cristián, V. y Sebastián, A. 2016. Effect of water availability on growth, water use efficiency and omega 3 (ALA) content in two phenotypes of chía (Salvia hispanica L.) established in the arid Mediterranean zone of Chile. Agricultural Water Management. 173:67-75. Doi:10.1016/j.agwat.2016.04.028.
Hoyle, G. L.; Steadman K. J.; Daws M. I. and Adkins S. W. 2008. Pre- and postharvest influences on seed dormancy status of an Australian goodeniaceae species, Goodenia fascicularis. Annals Bot. 102(1):93-101.
Ichihashi, Y.; Sinha, N. R. 2014. From genome to phenome and back in tomato. Curr. Opinion Plant Biol. 18:9-15.
ISTA. 2005. International Seed Testing Association. International Rules for Seed Testing. Zurich, Switzerland. 243 p.
Jamboonsri, W. 2012. Extending the range of an ancient crop, Salvia hispánica L. Genetic Res. Crop Evol. 171-178 pp.
Kobus-Cisowska, J.; Szymanowska, D.; Maciejewska, P.; Kmiecik, D.; Gramza-Michałowska, A.; Kulczyński, B. and Cielecka-Piontek, J. 2019. In vitro screening for acetylcholinesterase and butyrylcholinesterase inhibition and antimicrobial activity of chía seeds (Salvia hispanica). Elec. J. Biotechnol. 37:1-10. Doi: 10.1016/j.ejbt.2018.10.002.
Kromidas, S. 2006. HPLC made to measure: a practical handbook for optimization. Wiley-VCH, 2006. 754 p.
Lobo, M.; Delgado, O.; Cartagena, J. R.; Fernández, E. y Medina, C. I. 2007. Categorización de la germinación y la latencia en semillas de chirimoya (Annona cherimola L.) y guanábana (Annona muricata L.) como apoyo a programas de conservación de germoplasma. Agron. Colomb. 25(2):231-244.
Marineli, R. da S.; Moura, C. S.; Moraes, É. A.; Lenquiste, S. A.; Lollo, P. C. B.; Morato, P. N. and Maróstica, M. R. 2015. Chía (Salvia hispanica L.) enhances HSP, PGC-1α expressions and improves glucose tolerance in diet-induced obese rats. Nutrition. 31(5):740-748.
Medina-Santos, C. L.; Covarrubias-Prieto, J.; Aguirre-Mancilla, C. L.; Iturriaga, G.; Ramírez-Pimentel, J. G. y Raya-Pérez J. C. 2017. Evaluación de algunos compuestos nutraceúticos de la harina de chía. Ciencia y Tecnología Agropecuaria de México. 5(1):12-18.
Muñoz-Máximo, T.; Ocampo-Fletes, I.; Parra-Inzunza, F.; Cervantes-Vargas, J.; Argumedo-Macías, A. and Cruz-Ramírez, S. 2017. Proceso de producción y mecanismos de comercialización de chía (Salvia hispánica L.) por familias campesinas de los municipios de Atzitzihuacán y Tochimilco, Puebla, México. Nova Scientia. 9(19):788-818. doi: 10.21640/ns.v9i19.1104.
Palma-Rojas, C.; González, C.; Carrasco, B.; Silva, H.; & Silva-Robledo, H. 2017. Genetic, cytological and molecular characterization of chía (Salvia hispanica L.) provenances. Bio. System. Ecol. 73:16-21. doi.: 10.1016/j.bse.2017.05.003.
Puttini M.; Vuarant, C. M.; Fournier J.; Lesa, C.; Huter, C.; Ruíz-Díaz, J.; Romero, A. 2005. Determinación de ácidos oleico y linoleico del suero sanguíneo humano por HPLC de fase inversa. Ciencia, Docencia y Tecnología 16(30):179-192.
Seyyedi, S. M.; Tavakkol Afshari, R.; and Daneshmandi, M. S. 2018. The relationships between fatty acids and heterotrophic seedling growth in winter canola cultivars during accelerated seed aging process. South Afr. J. Bot. 119:353–361. Doi: 10.1016/j.sajb.2018.09.034.
Sreeedhar, R V.; Kumari; P. S. D. Rupwate; R. Rajasekharan; M. Srinivasan. 2015. Exploring triacylglycerol biosynthetic pathway in developing seeds of chía (Salvia hispanica L.): a transcriptomic approach. Plos One 10(4):e0123580.doi:10.1371/journal.pone.0123580.
Sun, Y.; X. Yan, S.; Zhang, N. and Wang. 2017. Grain yield and associated photosynthesis characteristics during dryland winter wheat cultivar replacement since 1940 on the Loess Plateau as affected by seedling rate. Emyrates J. Food Agric. 29(1):51-58.
Tenorio-Galindo, G.; Rodríguez-Trejo, D. A.; López-Ríos, G. 2008. Efecto del tamaño y color de la semilla en la germinación de Cecropia dotusifolia Bertol (Cecropiaceae). Agrociencia. 42(5):585-593.
Torossi, F. D. 2006. Reacciones en contexto: la transesterificación en la producción de biodiesel a partir de aceite de fritura usado. Anales de la Real Sociedad Española de Química. 102(3):43-49.
Vázquez-Carrillo, M. G.; Santiago-Ramos, D.; Salinas-Moreno, Y.; Rosas-Martínez, I.; Arellano-Vázquez, J. L.; Velázquez-Cardelas, G. A. y Espinoza-Calderón, A. 2012. Interacción genotipo-ambiente del rendimiento y calidad de grano y tortilla de híbridos de maíz en valles altos de Tlaxcala, México. Rev. Fitotec. Mex. 35(3):229-37.
Win, A. N.; Xue, Y.; Chen, B.; Liao, F.; Chen, F.; Yin, N. and Chai, Y. 2018. Chía (Salvia hispanica) experiment at a 30˚ N site in Sichuan Basin, China. Ciencia Rural. 48(9):1-12.
Xingú-López, A.; González-Huerta, A.; de la Cruz-Torres, E.; Sangerman-Jarquin, D. M., Orozco-de Rosas, G. y Rubí-Arriaga, M. 2017. Chía (Salvia hispánica L.) situación actual y tendencias futuras. Rev. Mex. Cienc. Agríc. 8(7):1619-1631.