Revista Mexicana Ciencias Agrícolas volume 10 number 5 June 30 - August 13, 2019
DOI: https://doi.org/10.29312/remexca.v10i5.1608
Article
Inoculation of arbuscular mycorrhizal fungi in the cultivation
of Heliconia stricta
Alma Gloria Uc-Ku1
Jesús Arreola-Enríquez1
Eugenio Carrillo-Avila1
Ma. Mónica Osnaya-González1
Alejandro Alarcón2
Ronald Ferrera-Cerrato2
Cesáreo Landeros-Sánchez3§
1Postgraduate College-Campus Campeche. Haltunchen-Edzná road km 17.5, Sihochac, Champotón, Campeche, Mexico. CP. 24450. (ucku@colpos.mx; jarreola@colpos.mx; ceugenio@colpos.mx; osnaya@colpos.mx). 2Postgraduate College-Campus Montecillo. Mexico-Texcoco highway km 36.5, Montecillo, Texcoco, State of Mexico, Mexico. CP. 56230. (alexala@colpos.mx; ronaldfc@colpos.mx). 3Postgraduate College-Veracruz Campus. Federal Highway Xalapa-Veracruz km 88.5, via Paso de Ovejas, between Puente Jula and Paso San Juan, Tepetates, Veracruz. CP. 91690.
§Corresponding author: clandero@colpos.mx.
Abstract
The inoculation with arbuscular mycorrhizal fungi has allowed a controlled development and growth in horticultural species; however, its effect has not been evaluated in some tropical ornamental crops such as heliconias. The objective was to evaluate the application of arbuscular mycorrhizal fungi (AMF) in the development of Heliconia stricta, taking as a general hypothesis that the use of AMF increases the absorption of nutrients such as nitrogen, phosphorus, potassium and favor their acclimatization and growth. The physiological development of H. stricta plants was evaluated in response to their inoculation with four AMF: Rhizophagus intraradices, Zac-19, mixture of strains and native inoculum in Mexico. The height of the plants, number of buds, photosynthetic rate with an IRGA photosynthesis analyzer, LI-COR 6400XT and mycorrhizal colonization by the Phillips and Hayman method were determined. The results showed statistically significant increases due to the effect of the non-native inoculum, on the height of the plant, the number of buds per plant, the photosynthetic rate and mycorrhizal colonization (p″ 0.05). The native inoculum produced statistically significant effects on the colonization of the roots (p″ 0.05). It is concluded that there is a statistically significant beneficial effect (p″ 0.05) of the mycorrhizal inoculation on the growth of the plants due to its higher height, number of buds per plant and photosynthetic rate.
Keywords: Heliconia stricta, inoculum, mycorrhizal colonization, photosynthetic rate.
Reception date: April 2019
Acceptance date: July 201
Introduction
The genus Heliconia belonging to the family Heliconiaceae is Neotropical, with approximately 200 known species, several of them ornamental due to the striking of their inflorescences (Calderón, 2009). Heliconias are monocotyledonous plants, with a rhizomatous growth that emit buds or offshoots. The height of these plants is between 0.45 m to 10 m. They grow in humid, subhumid, tropical and subtropical areas or with an irrigation system (Roca et al., 1991). The optimum temperature for its development is 28 °C with a range between 25 and 32 °C, cannot withstand frosts and does not produce flowers when they exceed 35 °C.
These flowers are not affected by the photoperiod and their flowering depends on the temperature. They should be planted in full sun or in areas where most of the day are illuminated by sunlight. Irrigation can help to solve the water deficit in areas with medium or low rainfall at the request (FHIA, 1995). The durability of the inflorescence in postharvest varies between the species and cultivars, on average it reaches 14 days. In the plant it can have a life of one to ten months, depending on the species, size and number of bracts (Maza et al., 2004). Heliconia stricta Huber, commonly known as platanillo, represents a growing alternative for producers in humid tropical zones. Like the banana plant, they have a pseudostem, made of petiole sheaths that overlap to support the weight of the flowers and leaves.
They emit flowers generally from April to December in the Northern Hemisphere, approximately two months after the bud of the pseudostem, depending mainly on the temperature and humidity conditions, when the pseudostem reaches a height of about one meter (Plant care today, 2019a). Each of the pseudostems blooms once and when the flowering is complete it collapses, dries and dies. The leaves of the plant resemble those of the plane trees, with bracts of flowers that emerge from the plant like a bunch of these fruits.
The flowers, large, succulent and long-lasting, have shades of red, orange, yellow, pink and green and have a shelf life as flower cut from 10 to 14 days if kept in water, although they are also marketed as pot plants or garden, in tropical or semi-tropical areas such as Florida, or even indoors. The ears of the flowers can be up to one meter long (Plant care today, 2019b).
Heliconias have gained great importance in the economic sector, are little known species and their tropical flowers have potential for the florist (Jerez, 2007). In recent years there is a boom in the world with the consumption of tropical exotic plants, hence countries like Colombia, Ecuador and Costa Rica, exploit much marketing. Exports of heliconias are around 24 000 to 30 000 stems a year, depending on the varieties. Canada, the United States of America, the Netherlands and Germany are the countries of destination for these exports, constituting a sector with high potential to generate employment at the local and regional level (Díaz et al., 2006).
According to the economic importance of the ornamental plants, they have evaluated the application of mycorrhizal fungi to promote their acclimatization and as growth promoters (Oliveira et al., 2011). This is because mycorrhizae have been shown to have the capacity to provide important benefits to their hosts, such as increasing the absorption of nutrients such as
nitrogen, phosphorus and potassium (Bucking et al., 2012), as well as favoring tolerance to water stress (Meddich et al., 2015), to salinity (Oztekin et al., 2013) and protection against root diseases and nematodes (Wehner et al., 2010; Baum et al., 2015).
Previous studies in ornamental plants of commercial importance, belonging to the family Zingiberaceae such as Zingiber spectabile Griff and Alpinia purpurata (Vieill.) K. Schum. (Silva et al., 2006), Tapeinochilos ananassae (Hassk.) K. Schum. (Oliveira et al., 2011) and Heliconia psittacorum L. (Santos et al., 2000), have shown significant effects in mycorrhizal colonization, growth and nutrient extraction.
The microorganisms of the rhizosphere contribute to plant growth, increasing the availability of limiting nutrients such as phosphorus and nitrogen, as well as the composition and activity of the bacterial group, which is strongly influenced by the type of vegetation present in the soil (Pedraza et al., 2010). On the other hand, it has been observed that the mycorrhizal association increases the phosphorus content in leaves and roots of Poncirus trifoliata (L.) Raf. (Wu et al., 2011).
Arbuscular mycorrhizae are a symbiotic association between Glomeromycota phylum fungi and the root of most plant species. This association is characterized by an exchange of nutrients and the generation of various benefits for the plant (Ortiz et al., 2015), mainly in terrestrial ecosystems and soils in rehabilitation or restoration (Evas et al., 2012).
These microorganisms favor the absorption of nutrients in plants and have great potential in sustainable agriculture as organic fertilizers (Garzón-Gómez and Cuervo-Andrade, 2008). Higa and Parr (2013) mentioned that the use of beneficial microorganisms, in addition to helping in the fixation of nitrogen and decomposing waste and organic waste, increases the growth and yield of the crops.
Arbuscular mycorrhizal fungi (AMF), modify the morphology of the roots to improve the increase of water and mineral uptake (Abd-Allah et al., 2015), also help to induce plant defenses to improve nutrition (He et al., 2017). There are some AMF that tend to associate with some species of host plants because of their wide distribution and because they interact symbiotically with a large number of plants and predominate more in association with exotic plant species. An example of the above is the Camellia Japonica L. (Theaceae, Theales), an exotic Magnoliophyta plant that has more than 3 000 ornamental cultivars (Borriello et al., 2015).
On the other hand, the use of AMF can contribute to soil fertility through the formation of aggregates, thanks to its mycelium and organic compounds that make it up mainly as lipids (Aguilera-Gómez et al., 2007). Montaño et al. (2012) states that formations of recalcitrant fungal products contribute to carbon sequestration and therefore mitigate the emission of carbon dioxide into the atmosphere. It also mentions that they promote the adaptation and survival of host plants that contribute to attenuate abiotic stress.
The present experiment was based on the inoculation of Heliconia stricta plants with the AMF (Rhizophagus intraradices, consortium Zac-19, mixture of strains, Acaulospora sp., Glomus callosum and Glomus sp.). The response variables were height, number of buds and photosynthetic
rate of inoculated plants, as well as level of spore colonization, both in soil and roots. The objective of this research was to evaluate the physiological development of Heliconia stricta Huber plants when inoculated with four different AMF inocula, which were Rhizophagus intraradices, Zac-19 consortium, mixture of strains, and native inoculum (Acaulospora sp., Glomus callosum and Glomus sp.).
Materials and methods
Vegetal material
The present investigation was carried out in the Campus Campeche Postgraduate College, in Sihochac municipality of Champoton in the state of Campeche, Mexico. It is located at coordinates 19° 50’ 21’’ north latitude and 90° 58’ 61’’ west longitude, at an altitude of 20 meters above sea level. 75 rhizomes were obtained from a cultivar of Heliconia stricta, selecting only 50, each weighing 250 g, which were washed with potable water to remove the soil and remaining plant residues. The rhizomes were disinfested with 1% Captan® for 30 min and washed with drinking water. Plastic pots in black color were used for the sowing of the rhizomes with a capacity of 2 L.
These were disinfected with a 2% commercial NaClO solution for 10 min and washed with potable water. In each pot, 810 g of sterile organic substrate (Peat-Moss) were placed and a rhizome was planted at five centimeters depth. The plants were in these containers until 120 days after inoculation (DDI), where they were irrigated every third day and later, they were transplanted in seedbeds in a greenhouse, where they remained until 220 DDI.
Mycorrhizal inocula
The AMF used were: 1) Rhizophagus intraradices (447 spores in 20 g of sand, 1 spp); 2) Zac-19 consortium composed of Glomus albidum, Claroideoglomus claroideum and Rhizophagus diaphanus (89 spores in 20 g of sand, 3 spp); 3) mixture of strains of Rhizophagus fasciculatus, Funneliformis geosporum, Sclerocystis sinuosa, Glomus sp., Glomus aggregatum, Acaulospora scrobiculata, Claroideoglomus etunicatum and Sclerocystis clavispora (previously isolated from orchards of Citrus latifolia Tanaka, Citrus aurantium L. and Carica papaya L., from the states of Tabasco, Yucatan and Veracruz) (59 spores in 20 g of sand, 8 spp.), these three inocula are part of the AMF germplasm bank of the soil microbiology area of the Postgraduate School (CP) Montecillo Campus; and 4).
Native inoculum (Acaulospora sp., Glomus callosum and Glomus sp.), isolated from the rhizosphere of the Heliconia plants established in the experimental field of the Campeche campus of the CP (97% of root colonization, 579 spores in 20 g of soil, 3 spp.). The scientific names of the mentioned AMF species are in accordance with the classification proposed by Redecker et al. (2013). The non-native mycorrhizal inoculums and the natives were applied at a rate of 20 g of inoculum (constituted by spores, rootlets, hyphae and sand) per plantlet, directly in the root system, 30 days after emergence.
Growing conditions in soil
The physical-chemical conditions of the soil were the following: pH of 7.5, electrical conductivity (CE) of 1.79 dS m-1, content of organic matter (OM) of 1.27%, phosphorus (P) of 17.52 mg kg-1, potassium (K) of 1.84 cmol+ kg-1, nitrogen (N) of 0.06%, calcium (Ca) of 48.6 cmol+ kg-1 and magnesium (Mg) 8.4 cmol+ kg-1, with 74.8% clay, 15.5% silt and 9.7 % sand, and classification according to its texture: clay. Five seedbeds of 12 m long, 1.5 m wide and 0.3 m high were prepared under greenhouse conditions.
The soil was disinfected by covering it with double black 600 gauge polyethylene and applying the chemical fungicide busan 30w to inhibit the growth of the native mycorrhizae of the soil, according to the manufacturer’s instructions. It was left to rest for seven days; later it was removed and its aeration was allowed for three more days. After that time, the same soil was used for the transplant. The plants from the pots were transplanted 120 DDI of the AMF in the previously prepared beds, considering a depth of coverage of the rhizome with soil of 5 cm and a distance between plants of 30 cm. It was watered every third day with a drip irrigation system.
Response variables evaluated
In the pot stage the height (cm) of the plants was measured every 30 days up to 120 DDI, at which time the pot stage ended. Once the soil transplant was carried out, measurements of this variable were continued every 20, up to 220 DDI. The measurement of the height considered the length from the base of the plant to the apex of the same. In addition, the number of buds per plant was evaluated, from the transplant of plants in soil, with a frequency of every 20 days up to 220 DDI.
Measurement of the photosynthetic rate
In each sampling, carried out every 20 days, five plants per treatment were randomly chosen. After transplanting in greenhouse seed beds, one leaf was selected for each plant to determine the photosynthetic rate, with an IRGA photosynthesis analyzer, LI-COR 6400 XT (LI-COR, 2019).
Mycorrhizal colonization was estimated using the thinning and root staining procedure proposed by Phillips and Hayman (1970), with observation of the roots in the LEICA® optical microscope, CME 1349521X, expressed as colonization percentage.
Experimental design and statistical analysis
A completely randomized experimental design was used, which included five treatments consisting of four inocula of AMF and one control without inoculation, with ten repetitions per treatment. The data obtained in each of the study variables were systematized and subjected to an analysis of variance using the statistical program Statistical Analysis System (SAS®) version 2014. In the cases in which significant effects of the treatments were found, the average values by the multiple means comparison test proposed by Tukey (p= 0.05).
Results
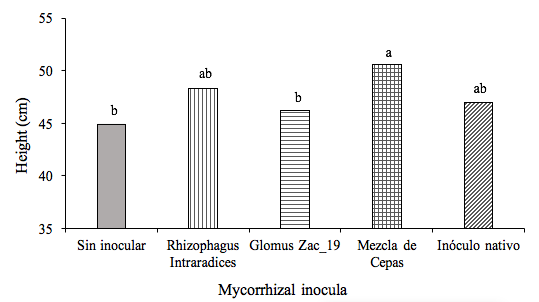
The inoculated plants, in general, had greater height with respect to the control during the whole crop, although statistically only the plants inoculated with the AMF strains mixture had higher height than the control at 220 DDI (Table 1) that corresponded with the point end of the experiment (Figure 1).
Figure 1. Effect of the inoculation of arbuscular mycorrhizal fungi on the height of Heliconia stricta plants, at 220 days after inoculation (DDI). Means followed by identical letters are statistically equal (Tukey, p= 0.05).
Table 1. Height of the plant (cm) in the samplings, by effect of mycorrhizal inoculation in Heliconia stricta plants.
Treatment | Days after mycorrhizal inoculation | |||||||||
0 | 30 | 60 | 90 | 120 | 140 | 160 | 180 | 200 | 220 | |
Without inoculating | 23.6a | 28.9b | 30.6b | 32.1b | 35.5b | 36.4c | 37.9c | 40.4c | 41.4c | 44.9b |
Rhizophagus Intraradices | 24.5a | 32.3ab | 35.3a | 37.7a | 39.1a | 40.1b | 41b | 42.3b | 44.2b | 48.3ab |
Glomus Zac-19 | 29.2a | 33.9a | 36.9a | 38.3a | 40.3a | 40.6ab | 42.1ab | 42.3b | 43.8b | 46.2b |
Mixture of strains | 25.6a | 34.8a | 37.2a | 38.9a | 41a | 42.2a | 42.9a | 44.2ª | 46.6a | 50.6a |
Native inoculum | 27a | 33.7a | 36a | 37.7a | 39.3a | 40.2ab | 41.2b | 42.1b | 43.1bc | 47ab |
DMS | 5.6 | 4.5 | 4.1 | 3.5 | 2 | 2 | 1.4 | 1.3 | 2.2 | 4.2 |
DMS= minimum significant difference; Tukey, p= 0.05. Means followed by the same letter in column are not statistically different.
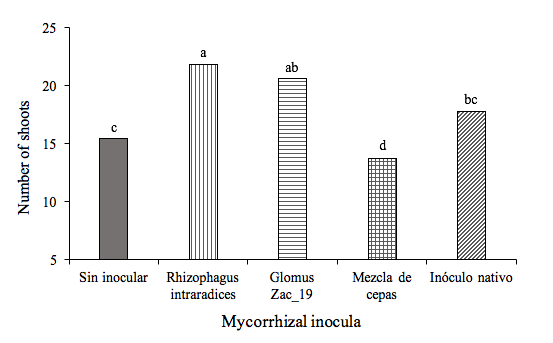
During the sampling period from 30 to 200 DDI there were no statistical differences for the number of buds per plant (Table 2). However, at 220 DDI the number of buds per plant was statistically different between treatments (Figure 2), which can be attributed to the effect of mycorrhizal inoculation. The treatment with R. intraradices presented statistically the highest number of buds per plant in relation to the other treatments and both the mixture of strains and the native inoculum, presented statistically the same number of buds as the control.
Figure 2. Effect of the inoculation of arbuscular mycorrhizal fungi on the number of buds per Heliconia stricta plant at 220 DDI. Means followed by identical letters are statistically equal (Tukey, p= 0.05).
Table 2. Number of buds per average plant in Heliconia stricta plants in the different samplings, due to mycorrhizal inoculation.
Treatment | Days after mycorrhizal inoculation | |||||
120 | 140 | 160 | 180 | 200 | 220 | |
Without inoculating | 1.7a | 4.2ª | 6.6ª | 8.8ab | 12.7bc | 15.4cd |
Rhizophagus intraradices | 2.1a | 5.3ª | 7.2ª | 10.3ab | 16.3a | 21.8a |
Glomus Zac-19 | 1.8a | 4.9ª | 7.4ª | 11a | 15.3ab | 20.6ab |
Mixture of strains | 1.6a | 4.2ª | 6.1ª | 8b | 11.9c | 13.7d |
Native inoculum | 1.5a | 3.9ª | 6.3ª | 8.4b | 14.1abc | 17.8bc |
DMS | 0.7 | 1.5 | 1.8 | 2.3 | 2.8 | 3.7 |
DMS= minimum significant difference; Tukey, p= 0.05. Means followed by the same letter in column are not statistically different.
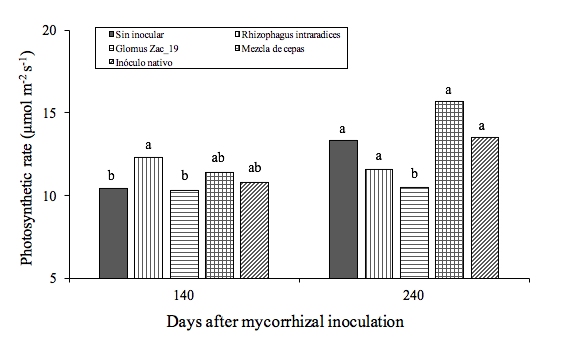
During the sampling period from 30 to 120 DDI there were no statistical differences for the photosynthetic rate of the plants. Mycorrhizal inoculation produced significant differences on this variable only from 140 DDI. The plants inoculated with Rhizophagus intraradices presented higher photosynthetic rate, with respect to the control, which was not observed for Zac-19, the mixture of strains and the native inoculum. This may be due to the fact that heliconia plants tend to associate more with strains of the Rhizophagus intraradices fungus than with the rest of the strains used in the work. This same strain produced a significantly higher height and number of buds than the rest.
At 240 DDI, the plants treated with the Zac-19 consortium showed statistically the lowest photosynthetic rate with respect to the other treatments, including the control (Figure 3, Table 3).
Figure 3. Effect of inoculation with mycorrhizal fungi on the photosynthetic rate (µmol m-2 s-1) in Heliconia stricta plants.
The means, followed by identical letters on the bars in their corresponding day of sampling, are statistically equal (Tukey, p= 0.05). Each bar corresponds to a different treatment: Control, Rhizophagus intraradices, consortium Zac-19, mixture of strains and native inoculum, at 140 (left) and 240 DDI (right).
Table 3. Average photosynthetic rate in plants (hour) of Heliconia stricta in the different samples by effect of inoculation.
Treatment | Days after mycorrhizal inoculation | |||||
120 | 140 | 160 | 180 | 200 | 240 | |
Without inoculating | 4.2a | 10.4b | 14.6a | 8.5a | 13.9a | 13.3a |
Rhizophagus intraradices | 1bc | 12.3a | 10.6a | 10.2a | 13.1a | 11.6a |
Glomus Zac-19 | 0.8c | 10.3b | 9.1a | 11.3a | 12.7a | 10.5b |
Mixture of strains | 1.1bc | 11.4ab | 10.5a | 10.7a | 14.6a | 15.7a |
Native inoculum | 1.7b | 10.8ab | 14.6a | 13a | 13.6a | 13.5a |
DMS (Tukey p= 0.05) | 1.5 | 0.8 | 6.4 | 7.6 | 4.9 | 5.2 |
DMS= minimum significant difference; Tukey, p= 0.05. Means followed by the same letter in column are not statistically different.
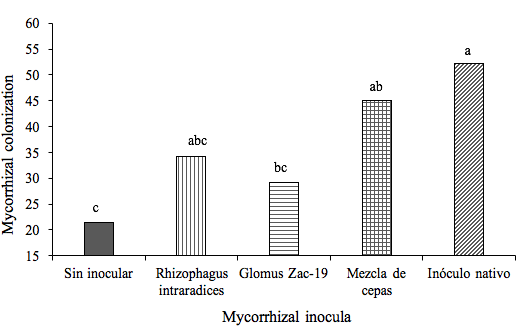
The mycorrhizal colonization of the root showed significant statistical differences (Tukey, p≤ 0.05) due to the effect of the AMF treatments only from the 120 DDI age of the crop, which could be explained by the time taken by the mycorrhizal fungi to infect the root of heliconia plants. The treatments with greater total mycorrhizal colonization, significantly different from the control, corresponded to the native inoculum and to the inoculation of the mixture of strains (Figure 4). It
was contemplated that the roots of the control showed a mycorrhizal colonization that appeared naturally without significant statistical differences with the colonization of the roots of the plants inoculated with R. intraradices and with the Zac-19 consortium (Table 4).
Figure 4. Effect of AMF-based inoculation on mycorrhizal colonization (number of average spores per root segment) of Heliconia stricta at 220 DDI of the culture. Means followed by identical letters are statistically equal (Tukey, p= 0.05).
Table 4. Mycorrhizal colonization (%) in root of Heliconia stricta plants in the different samples, by effect of inoculation.
Treatment | Days after mycorrhizal inoculation | |||
120 | 150 | 180 | 220 | |
Without inoculating | 0.1a | 1.2b | 7.2b | 21.5c |
Rhizophagus intraradices | 0.3a | 1.2b | 9.3ab | 34.3abc |
Glomus Zac-19 | 0.1a | 3.7b | 10.2ab | 29.1bc |
Mixture of strains | 0.3a | 13.3a | 25.8ª | 45ab |
Native inoculum | 0.3a | 5ab | 20.3ab | 52.2ª |
DMS | 0.4 | 9.2 | 16.6 | 18.3 |
DMS= minimum significant difference; Tukey, p= 0.05. Means followed by the same letter in column are not statistically different.
Discussion
The results obtained in the present work denote significant effects of the inoculation of plants with different strains of mycorrhizal fungi in most of the response variables evaluated. At the height of the plant, the mixture of AMF strains produced statistically higher values than the control treatment (p″ 0.05) at 220 DDI. Similar results have been observed with the inoculation of AMF in Calendula, where a relationship between root colonization and growth effect was studied (Vaingankar and Rodríguez, 2015).
In other studies, there was an increase in the growth and survival of C. papaya when they were inoculated with mycorrhizae of native consortia (Montaño et al., 2012). Vaingankar and Rodríguez (2015) also analyzed an increase in plant height in Tagetes erecta and Zinnia elegans with the inoculation of AMF. Vaingankar and Rodríguez (2015) demonstrated that the application of AMF had a significant effect on the height of the C. infundibuliformis plant. The treatment mix of strains, contains a greater number of species compared to the other treatments; therefore, it is attributed to that because it presented higher height, although less colonization was perceived in the roots.
This can be compared with studies carried out in orchid plants with respect to colonization, effects of inoculation and growth (Zhao et al., 2014) and in tomato with Glomus (Bañuelos et al., 2014). This means that it does not necessarily have to have a high colonization to observe the effect of the fungi on the plants, rather it will depend on the species of the fungus and the type of plant.
On the other hand, the non-inoculated plants showed a height statistically similar to the height of those that were inoculated with the rest of the treatments evaluated, in which colonization occurred, but no promotion of growth.
Although AMF are morphologically restricted to roots, colonization has been shown to beneficially affect roots and buds (Romero et al., 2017). This same behavior was observed in the present work, where there was an increase in the number of buds per plant in all the treatments, although only Rhizophagus intraradices and Zac-19 induced values significantly higher than the control at 220 DDI.
In other studies, it has been observed that Camellia Sinensis has shown colonization and shoot production (Borriello et al., 2015). Vaingankar and Rodríguez (2015) and Perner et al. (2007) reported an increase in the number of buds and flowers with AMF in Tagetes erecta, Zinnia elegans, Callistephus, Pelargonium and tomato. A significant finding of these studies for practical horticulture was the increase in the number of buds with inoculation with AMF. According to the commercial quality standards of the plant, a greater number of buds is desirable since each bud constitutes a potential flower of good quality if the management in terms of moisture and nutrition of the plant is adequate. In the present work the use of AMF produced a greater number of buds and a significantly higher plant height.
AMF often modify the gaseous exchange of the plant (Auge et al., 2016). The speed of photosynthesis, stomatal conductance (gs) and transpiration (E) are often higher in plants with AMF than in controls. In this study it was found that the photosynthetic rate at 140 DDI in the Rhizophagus intraradices treatment was significantly higher than with the Zac-19 consortium and with the control. These results are similar to that found in crops such as corn and rice by Andrade et al. (2015). The inoculation of AMF can improve the photosynthetic rate, the growth and the development of the plants (He et al., 2017).
The increased photosynthetic activity of plants with AMF could be due to non-stomatal factors such as higher chlorophyll content and rubisco activity (Porcel et al., 2015). Porcel et al. (2015) point out that fungi improve the exchange of carbohydrates and stimulate the rate of photosynthesis. The authors demonstrated the modulation of the efficiency of photosynthesis by the AMF symbiosis in rose, pistachio, poplar and citrus plants. The increase in the photosynthetic rate, observed in the present work with the use of AMF, leads to a reduction in the cycle of the plant, which in this case is beneficial to reduce the time to obtain flowers. However, as far as we know, there are no reports in the scientific literature that have evaluated this aspect, nor their effect on the size and quality of heliconia flowers.
Regarding mycorrhizal colonization in the root of H. stricta plants, significantly higher mycorrhization (p″ 0.05) was observed with the native inoculum and with the mixture of strains, at 220 DDI, with respect to that observed in the control. These results are similar to those found by Borriello et al. (2015), who inoculated AMF in exotic Camellia, finding high levels of mycorrhization.
In other studies it has been observed that rice plants are colonized by Claroideoglomus etunicatum (Porcel et al., 2015), which is comparable with the results found in the present work, both with the native inoculum and with the mixture of strains, since both colonized the roots of heliconia, highlighting the native inoculum. It should be mentioned that in other studies it has been observed that insects can influence the colonization of roots by AMF and therefore affect the growth of plants (He et al., 2017).
Conclusions
The cultivation of Heliconia stricta in the greenhouse was favored by mycorrhizal inoculation, which resulted in a better development of the plants, with a significantly higher growth, a greater number of buds and a statistically higher photosynthetic rate. A beneficial effect of mycorrhizal inoculation on the growth of Heliconia stricta plants was observed, which could be observed in the different evaluated study variables, which had a statistically superior behavior with the mixture of strains and native inoculum treatments. Such effects were translated into a better development of the plants, especially in the case of the native inoculum of the area.
Acknowledgments
We thank the Foundation Avanza and Pablo García, from the Government of the State of Campeche, for the scholarship granted to the first author for the completion of Master of Science studies.
Cited literature
Abd-Allah, E. F.; Abeer, H.; Alqarawi, A. A. and Alwathnani-Hend, A. 2015. Alleviation of adverse impact of cadmium stress in sunflower (Helianthus annuus L.) by arbuscular mycorrhizal fungi. Pak. J. Bot. 47(2):785-795.
Aguilera-Gómez, L. I.; Olalde-Portugal, V.; Arriaga, M. R. y Contreras-Alonso, R. 2007. Micorrizas arbusculares. Ciencia Ergo Sum. 14(3):300-306.
Andrade, S. A. L.; Domínguez, A. P. and Mazzafera, P. 2015. Photosynthesis is induced in rice plants that associate with arbuscular mycorrhizal fungi and are grown under arsenate and arsenite stress. Chemosphere. 134:141-149.
Augé, R. M.; Toler, H. D. and Saxton, A. M. 2016. Mycorrhizal stimulation of leaf gas exchange in relation to root colonization, shoot size, leaf phosphorus and nitrogen: a quantitative analysis of the literature using meta-regression. Front. Plant Sci. 7:1084. doi: 10.3389/fpls.2016.01084.
Banuelos, J.; Alarcón, A.; Larsen, J.; Cruz-Sánchez, S. and Trejo, D. 2014. Interactions between arbuscular mycorrhizal fungi and Meloidogyne incognita in the ornamental plant Impatiens balsamina. J. Soil Sci. Plant Nutr. 14(1):63-74.
Baum, C.; El-Tohamy, W. and Gruda, N. 2015. Increasing the productivity and product quality of vegetable crops using arbuscular mycorrhizal fungi: a review. Sci. Hortic. 187:131-141.
Borriello, R.; Berruti, A.; Lumini, E.; Della Beffa, M. T.; Scariot, V. and Bianciotto, V. 2015. Edaphic factors trigger diverse AM fungal communities associated to exotic camellias in closely located Lake Maggiore (Italy) sites. Mycorrhiza. 25(4):253-265.
Bücking, H.; Liepold, E. and Ambilwade, P. 2012. The role of the mycorrhizal symbiosis in nutrient uptake of plants and the regulatory mechanisms underlying these transport processes. In: Plant Sci. Kumar, D. N. and Charan, S. S. (Ed.). InTech. Doi: 10.5772/52570.
Calderón de Rzedowski, G. 2009. Heliconiaceae. Flora del bajío y de regiones adyacentes. Instituto Nacional de Ecología. Centro Regional del Bajío, Pátzcuaro, México. 161:1-7.
Díaz, M. J. 2006. Diagnóstico de la cadena productiva de Heliconias y follajes en los departamentos del eje cafetero y Valle del Cauca (Colombia). Biocomercio Sostenible. United Nations.
Evas, L. V.; Álvarez-Sánchez, J.; González-Chávez, M. D. C. A. and Maldonado-Mendoza, I. E. 2012. Research on arbuscular mycorrhizae in Mexico: an historical synthesis and future prospects. Symbiosis. 57(3):111-126.
FHIA. 1995. Fundación Hondureña de Investigación Agrícola. Introducción a ornamentales tropicales. Honduras. 131 p.
Garzón-Gómez, J. P. y Cuervo-Andrade, J. L. 2008. Producción de Pleurotus ostreatus sobre residuos sólidos lignocelulósicos de diferente procedencia. Nova. 6(10):101-236.
He, L.; Li, C. and Liu, R. 2017. Indirect interactions between arbuscular mycorrhizal fungi and Spodoptera exigua alter photosynthesis and plant endogenous hormones. Mycorrhiza. 27(6):1-11.
Higa, T. y Parr, J. F. 2013. Microorganismos benéficos y efectivos para una agricultura y medio ambiente sostenibles. Maryland (USA): Centro internacional de Investigación de Agricultura Natural, Departamento de Agricultura de los Estados Unidos. 13 p.
Jerez, E. 2007. El cultivo de las Heliconias. Cultivos Tropicales. 28(1):29-35.
LI-COR. 2019. LI-COR® Instrumentation for biological sciences. https://www.licor.com/ env/products/photosynthesis/LI-6400XT/.
Maza, V. 2004. Cultivo, cosecha y postcosecha de Heliconias y flores de corte. Primera edición. Jardín Botánico. 193 p.
Meddich, A.; Jaiti, F.; Bourzik, W.; El Asli, A. and Hafidi, M. 2015. Use of mycorrhizal fungi as a strategy for improving the drought tolerance in date palm (Phoenix dactylifera). Sci. Hortic. 192:468-474.
Montaño, N. M.; Alarcón, A.; Camargo-Ricalde, S. L.; Hernández-Cuevas, L. V.; Álvarez-Sánchez, J.; González-Chávez, M. D. C. A. and Maldonado-Mendoza, I. E. 2012. Research on arbuscular mycorrhizae in Mexico: an historical synthesis and future prospects. Symbiosis. 57(3):111-126.
Oliveira, J. R. G. D.; Morais, T. A. D. L.; Melo, N. F. D. and Yano-Melo, A. M. 2011. Acclimatization of Tapeinochilos ananassae plantlets in association with arbuscular mycorrhizal fungi. Pesquisa Agropecuária Brasileira. 46(9):1099-1104.
Ortíz-Acevedo, A.; Osorio-Vega, N. W.; Echeverri-Gómez, J.; González-Murillo, O. A. y Medina-Sierra, M. 2015. Fisiología de los hongos formadores de micorrizas arbusculares. Livestock Res. Rural Develop. 27:188. http://www.lrrd.org/lrrd27/9/orti27188.html.
Oztekin, G. B.; Tuzel, Y. and Tuzel, I. H. 2013. Does mycorrhiza improve salinity tolerance in grafted plants? Sci. Hortic. 149:55-60.
Pedraza, R. O.; Teixeira, K. R.; Fernández Scavino, A.; García de Salamone, I.; Baca, B. E.; Azcón, R. y Bonilla, R. 2010. Microorganismos que mejoran el crecimiento de las plantas y la calidad de los suelos. Revisión. Corpoica. Ciencia y Tecnología Agropecuaria. 11(2):155-164.
Perner, H.; Schwarz, D.; Bruns, C.; Mäder, P. and George, E. 2007. Effect of arbuscular mycorrhizal colonization and two levels of compost supply on nutrient uptake and flowering of pelargonium plants. Mycorrhiza. 17(5):469-474.
Phillips, J. M. and Hayman, D. S. 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of the British Mycological Society. 55(1):158-161.
Plant Care Today. 2019b. Heliconia stricta flowers perfect for containers. https://plantcaretoday. com/heliconia-stricta-flowers-perfect-containers.html.
Plant Care Today. 2019a. Growing heliconia plants: how to care for the lobster claw flower. https://plantcaretoday.com/heliconia-plant-care.html.
Porcel, R.; Redondo-Gómez, S.; Mateos-Naranjo, E.; Aroca, R.; García, R. and Ruiz-Lozano, J. M. 2015. Arbuscular mycorrhizal symbiosis ameliorates the optimum quantum yield of photosystem II and reduces non-photochemical quenching in rice plants subjected to salt stress. J. Plant Physiol. 185:75-83.
Roca, W. M. y Mroginski, L. A. 1991. CIAT. Cultivo de tejidos vegetales: fundamentos y aplicaciones. Cali, Colombia. 970 p.
Romero‐Munar, A.; Del‐Saz, N. F.; Ribas‐Carbó, M.; Flexas, J.; Baraza, E.; Florez‐Sarasa, I. and Gulías, J. 2017. Arbuscular mycorrhizal symbiosis with Arundo donax decreases root respiration and increases both photosynthesis and plant biomass accumulation. Plant, Cell Environ. 40(7):1115-1126.
Santos, B. A.; Silva, G. A.; Maia, L. C. and Alves, M. V. 2000. Mycorrhizae in Monocotyledonae of Northeast Brazil: subclasses Alismatidae, Arecidae and Zingiberidae. Mycorrhiza. 10(3):151-153.
Silva, M. A. D.; Silva, F. S. B. D.; Yano-Melo, A. M.; Melo, N. F. D. and Maia, L. C. 2006. Mycorrhizal arbuscular and vermicompost fungus in the acclimatization of purple Alpinia (Viell.) Schum e Zingiber spectabile Griff. (Zingiberaceae). Acta Botânica Brasilica. 20(2):249-256.
Tukey, J. W. 1991. The philosophy of multiple comparisons. Statistical science. 6(1):100-116.
Vaingankar, J. D. and Rodríguez, B. F. 2015. Effect of arbuscular mycorrhizal (AM) inoculation on growth and flowering in Crossandra infundibuliformis (L.) Nees. J. Plant Nutr. 38(10):1478-1488.
Wehner, J.; Antunes, P. M.; Powell, J. R.; Mazukatow, J. and Rillig, M. C. 2010. Plant pathogen protection by arbuscular mycorrhizas: a role for fungal diversity? Pedobiologia. 53(3):197-201.
Wu, Q. S.; Zou, N. and He, X. H. 2011. Differences of hyphal and soil phosphatase activities in drought-stressed mycorrhizal trifoliate orange (Poncirus trifoliata) seedlings. Sci. Hortic. 129(2):294-298.
Zhao, X. L.; Yang, J. Z.; Liu, S.; Chen, C. L.; Zhu, H. Y. and Cao, J. X. 2014. The colonization patterns of different fungi on roots of Cymbidium hybridum plantlets and their respective inoculation effects on growth and nutrient uptake of orchid plantlets. World J. Microbiol. Biotechnol. 30(7):1993-2003.